Асимметрия в симметрии

В-четвертых, при погружении стимулирующего и регистрирующего электродов в кору противоположных полушарий для анализа послойной организации каллозальных проекций было обнаружено, что при стимуляции на глубине 1 мм максимум транскаллозального ответа наблюдался в противоположном полушарии на глубине 2 мм, а при стимуляции на глубине 2 мм такой максимум отмечался на глубине 1 мм. С одной стороны… Читать ещё >
Асимметрия в симметрии (реферат, курсовая, диплом, контрольная)
В середине семидесятых годов прошлого века В. Л. Бианки была разработана специальная методика множественной топографической регистрации вызванных потенциалов. Ее особенность заключалась в последовательной регистрации прямых и транскаллозальных ответов во многих точках поверхности коры, располагающихся на равном расстоянии друг от друга. Прямые корковые ответы — это вызванные потенциалы, регистрируемые в том же полушарии, которое подвергнуто стимуляции непосредственно или через систему афферентных волокон. Транскаллозальные ответы — это вызванные потенциалы, которые можно зарегистрировать в коре одного полушария при электрической стимуляции симметричного пункта другого полушария. На рисунке 42 представлены примеры транскаллозальных ответов, зарегистрированных у кошки при электрической стимуляции сенсомоторной коры.
На рисунке показаны транскаллозальные ответы (то есть ответы, возникающие в полушарии, противоположном раздражаемому), зарегистрированные в разных слоях сенсомоторной коры кошки. Цифры слева — глубина погружения регистрирующего электрода в кору мозга. Цифры внизу — калибровка времени (в миллисекундах) и амплитуды ответа (в милливольтах).
Благодаря этой методике исследователи получили несколько фактов, свидетельствующих о том, что хотя морфологически полушария и объединены между собой в основном гомотопическими связями, передача по этим связям возбуждающих и тормозящих влияний от нейронов одного полушария к нейронам другого не всегда строго гомотопична. Л. В. Бианки выделяет пять таких фактов.
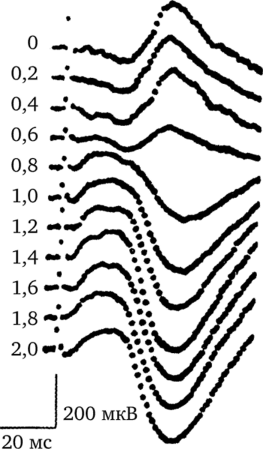
Рис. 42. Примеры транскаллозальных ответов, зарегистрированных в сенсомоторной коре кошки1
Вызванные потенциалы — это суммарный ответ клеток мозга на раздражение рецептивных полей, афферентных волокон или непосредственно каких-либо структур ЦНС стимулами различной модальности. Раздражение может быть механическим, световым, звуковым, электрическим и так далее в зависимости от того, какую структуру и с какой целью раздражает экспериментатор.
На это раздражение в нервной системе после небольшого латентного периода (у животных от 2 до 8 мс) возникает ответ — вызванный потенциал, состоящий из негативных и позитивных компонентов.
Во-первых, согласно правилу гомотопичности, транскаллозальные ответы должны регистрироваться на поверхности коры площадью не более 4 мм2. Именно такая площадь соответствует предполагаемой зоне терминальных ветвлений гомотопического каллозального волокна и его связей с нейронами-мишенями в противоположном полушарии (X. Чанг, 1953 г.; Ф. Бремер, 1958 г.). Однако результаты исследований показали, что локальная стимуляция коры одного полушария вызывает активацию значительной зоны коры противоположного полушария, площадь которой намного превышает ожидаемые 4 мм2. То есть раздражение одной точки в стимулируемом полушарии каким-то образом вызывает активацию множества точек в противоположной гемисфере.
Во-вторых, при изменении местоположения стимулирующего электрода и при неподвижном регистрирующем электроде было установлено, что на активность тех точек, которые расположены под регистрирующим электродом, можно влиять, раздражая различные, достаточно удаленные друг от друга пункты противоположного полушария. Это свидетельствовало о том, что каллозальные волокна приходят к исследуемой точке от значительной площади раздражаемого полушария. Не от одной точки к другой, что соответствовало бы принципу гомотопичности, а от нескольких точек к одной. При изменении положения регистрирующего электрода смещался центр (фокус) зоны активации, изменялась его ориентация и форма.
В-третьих, было выявлено, что на один и тот же стимул зона прямых корковых ответов (то есть вызванных ответов, регистрируемых в том же полушарии, которое подвергнуто стимуляции) всегда оказывается более широкой, чем зона транскаллозальных ответов (то есть вызванных ответов, регистрируемых в противоположном полушарии). Зона прямых корковых ответов имела форму кольца, а очаг транскаллозальных ответов мог стягиваться в точку. Получалось некоторое подобие воронки, пронизывающей мозолистое тело. Раструб этой воронки, воспринимающий раздражение, находился в том полушарии, которое раздражали, а узенькое устье — в противоположном. Активность как бы стекала с больших площадей одного полушария в определенное место другого. Это тоже не очень соответствовало принципу гомотопичности.
В-четвертых, при погружении стимулирующего и регистрирующего электродов в кору противоположных полушарий для анализа послойной организации каллозальных проекций было обнаружено, что при стимуляции на глубине 1 мм максимум транскаллозального ответа наблюдался в противоположном полушарии на глубине 2 мм, а при стимуляции на глубине 2 мм такой максимум отмечался на глубине 1 мм. С одной стороны, результаты этого эксперимента подтвердили данные об организации каллозальных проекций по принципу «слой в слой», постулированному в свое время американскими исследователями Якобсоном и Трояновским (1974 г.). С другой стороны, они показали, что возможно в коре существуют и перекрещивающиеся гетеротопические межполушарные пути, организованные по принципу «один слой — другой слой» и направленные от третьего слоя к пятому и от пятого к третьему.
И наконец, в-пятых, было показано, что фокус максимальной активности транскаллозального ответа не всегда строго симметричен точке раздражения. Фокус максимальной активности — это та точка в коре полушария, противоположного раздражению, в которой транскаллозальные ответы регистрируются с наиболее высокой амплитудой и наиболее однообразной формой. Эксперименты показали, что при стимуляции фокус максимальной активности мог быть смещен в оральнокаудальном направлении на 1,4—2,7 мм относительно места, строго симметричного точке раздражения. Вот это-то смещение фокуса максимальной активности транскаллозального ответа относительно точки раздражения противоположного полушария и стало краеугольным камнем построения рассуждений о том, почему симметричность анатомических связей между полушариями мозга может лежать в основе функциональной асимметрии (рис. 43).
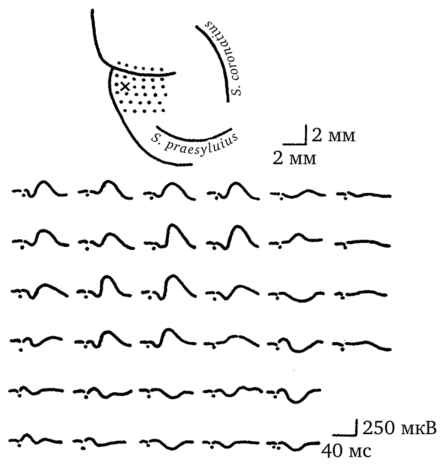
Рис. 43. Смещение фокуса максимальной активности транскаллозальных ответов, зарегистрированных в сенсомоторной коре кошки1
В верхней части рисунка — участок полушария, контралатерального тому месту, на который наносилось раздражение. Точками помечены места регистрации транскаллозальных потенциалов. Крестиком помечена точка, симметричная месту раздражения.
В нижней части рисунка — транскаллозальные ответы, представленные в соответствии с расположением точек в верхней части слайда. Хорошо видно, что фокус максимальной активности транскаллозального ответа смещен вправо относительно точки, симметричной месту раздражения.
В правом верхнем углу — масштаб. В правом нижнем — калибровка времени и амплитуды Для объяснения причин смещения фокуса максимальной активности транскаллозального ответа были выдвинуты два предположения. Согласно первому предположению такое смещение могло быть обусловлено преобладанием гетеротопических транскаллозальных связей над гомотопическими. Но если согласиться с этим предположением, то тогда нужно признать, что основной принцип организации каллозальных связей — принцип гомотопичности — не верен. А этот принцип хорошо обоснован и подкреплен многочисленными экспериментальными данными. Согласно второму предположению, смещение фокуса максимальной активности определяется суммацией возбуждения, распро1 В. Л. Бианки, 1989 г.
страняющегося как по транскаллозальным, так и (что особенно важно) по внутриполушарным путям. Это предположение не вступало в конфликт с основным принципом, но так же, как и первое, требовало доказательств.
Для проверки этих предположений были проведены эксперименты, в которых перерезали мозолистое тело, а также обрезали кору вокруг точек стимуляции и вокруг симметричных им участков в противоположном полушарии. После рассечения мозолистого тела и обрезки коры вокруг точки раздражения вызванные потенциалы в контралатеральном полушарии в точке, геометрически симметричной месту раздражения, не регистрировались. Это свидетельствовало о том, что возникновение этих потенциалов обусловлено главным образом распространением возбуждения по гомотопическим волокнам из точки раздражения в точку регистрации.
Таким образом, первое предположение оказалось несостоятельным. Раз транскаллозальные ответы пропадают в результате пересечения мозолистого тела и изоляции точки раздражения, значит их возникновение обусловлено приходом возбуждения по гомотопическим (симметричным) каллозальным волокнам, поскольку все гетеротопические пути распространения возбуждения в полном смысле этого слова отрезаны из-за хирургического вмешательства в область стимуляции. Но почему эти ответы максимальны в точке, не строго симметричной раздражению? Что приводит к их усилению в стороне от этой точки?
В. Л. Бианки предположил, что это связано с суммацией возбуждения, приходящего по межполушарным волокнам, с возбуждением, приходящим по ассоциативным волокнам воспринимающего полушария. То, что внутриполушарные связи воспринимающей гемисферы участвуют в создании фокуса максимальной активности, доказывают опыты с обрезкой коры вокруг точки, симметричной раздражению. После такой операции в том месте, где ранее наблюдался фокус максимальной активности, ответы вообще не возникают. А это можно объяснить только тем, что в результате хирургической процедуры исчезли пути, по которым возбуждение, пришедшее из одного полушария в другое, могло бы достигнуть этого места. А достигнуть его оно могло только по внутриполушарным волокнам. И Бианки предположил, что в фокусе максимальной активности преобладает внутриполушарный поток возбуждения, а в точке, симметричной раздражению, — транскаллозальный. Именно внутриполушарные волокна обеспечивают смещение фокуса в сторону от точки, в которую пришло возбуждение. И, поскольку согласно данным О. С. Адрианова (1976 г.) ассоциативные волокна ориентированы преимущественно в орально-каудальном направлении, фокус максимальной активности смещается относительно точки, симметричной раздражению, именно в этом направлении в ту или иную сторону. Это очень важный момент, поскольку он свидетельствует о том, что функциональная асимметрия возникает на основе морфологически симметричных структур. Если бы описанное смещение фокуса максимальной активности транскаллозального ответа зависело от морфологической несимметричности полушарий, то это смещение можно было бы наблюдать в любом направлении, а не только в орально-каудальном.
Таким образом, В. Л. Бианки и его сотрудниками было показано, что гомотопические потоки транскаллозального возбуждения становятся асимметричными благодаря внутриполушарным влияниям. По его собственному, очень образному выражению «…гомотопика является как бы той канвой, на которой интракортикальные связи и гетеротопические каллозальные проекции вышивают свой асимметричный узор». Из этого следует, что основой функциональной асимметрии больших полушарий головного мозга является гомотопичность межполушарных связей, то есть их симметрия.