Стехиометрия ENaC и модель поры

Рис. 10. Предсказанная структура поры и области короны ENaC. Предсказанные порообразующие трансмембранные домены гетеротримерного ENaC параллельно плазматической мембране (А). Консервативный Trp52 на внутриклеточный N-конце TM1 в-hENaC изображен с голубой боковой цепью. Консервативный Tyr (358 и 391) в предполагаемых петлях связи ви б-ENaC, которые находятся чуть выше поры, изображены с синими… Читать ещё >
Стехиометрия ENaC и модель поры (реферат, курсовая, диплом, контрольная)
ENaC представляет собой гетеромультимер состоящий из трех гомологичных субъединиц: б, в и г (Canessa C. M et al., 1994). Вопрос о стехиометрическом соотношении субъединиц встал сразу же после их первоначального клонирования. Результаты и выводы по данному вопросу были неоднозначными и предполагали структуры от тетрамерной (2б, в, г) до крупных комплексов, содержащих по две и более субъединицы каждого вида. Долгое время наиболее распространенной моделью строения ENaC была тетрамерная модель (Рис. 6). Исследования по данной проблеме были основаны на оценке стехиометрии путем смешивания мутантных субъединиц и субъединиц дикого типа, предполагая, что эти субъединицы могут свободно ассоциировать друг с другом и образовывать различные комбинации (Firsov D. et al., 1998; Kosari F. et al., 1998; Eskandari S. et al., 1999; Anantharam A. et al., 2007). Эти данные дополнялись также и другими методами: градиентом осаждения сахарозы (Snyder P.M. et al., 1998; Dijkink L. et al., 2002), поверхостной экспрессией (Firsov D. et al., 1998), электронной микроскопией (Eskandari S. et al., 1999), флуоресцентным анализом (Staruschenko A. et al., 2005). Однако после открытия гетеротримерной модели каналов ASIC, относящихся к тому же суперсемейству Deg/ENaC, появились идеи об аналогичном гетеротримерном строении и ENaC (Canessa C.M., 2007) (Рис. 7). Рассмотрение тримерной структуры ENaC породило вопрос о круговой ориентации субъединиц б-в-г или б-г-в (по часовой стрелке с внешней стороны мембраны) (Рис. 8). На основе результатов эксперимента построенного на Cl—зависимом ингибировании ENaC, предполагается, что ENaC является б-г-в тримером (Collier D.M., Snyder P.M., 2010). Эту идею подтверждает и другое исследование, основанное на связывании двухвалентных катионов Cu2+ с внеклеточными доменами ENaC (Chen J. et al., 2011). Однако не исключено существование популяций ENaC с расположением субъединиц б-в-г (Рис. 8).
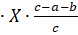
Рис. 7. Модель тримерной организации субъединиц ENaC. (Модифицировано из Bhalla, Hallows, 2008)
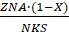
Рис. 8. Возможные варианты сборки гетеротримера ENaC бгв (A), бвг (B). Короткий пунктир — область домена thumb («большой палец»); длинный пунктир — область домена palm («ладонь»). (модифицировано из Collier, Snyder, 2011)
Предсказанная по аналогии с ASIC гетеротримерная модель ENaC представляет собой бокалообразную структуру с крупным воронковидным внеклеточным участком, узким трансмембранным сегментом и расширением в области концевых цитоплазматических доменов. Внешняя воронка представлена экстраклеточными доменами и C-концевым участками сегментов TM1. Непосредственно пора и селективный фильтр образованы сегментами TM2 (Рис. 9). Внутриклеточная часть сложена Nи С-концевыми доменами (Stockand J.D. et al., 2008). На внеклеточном конце сегментов TM2, в области наружного устья канала имеются высококонсервативные остатки тирозина, которые, по-видимому играют роль в воротных характеристиках канала. На внутриклеточном конце сегментов TM1 обнаружены высококонсервативные остатки триптофана, выдающиеся своими радикалами в просвет поры, мутации в которых предположительно должны были менять воротные характеристики канала, однако, исследование по данному вопросу не показало изменений в воротных характеристиках, при этом обнаружено влияние на амилорид-чувствительные токи и нарушение поверхностной экспрессии канала, возможно, из-за нарушения сборки канала (Kashlan O.B. et al., 2006).
Селективный фильтр поры ENaC позволяет проникать через канал катионам Na+ и более мелким катионам Li+ и H+, при этом более крупные катионы, такие как катионы K+, Rb+, Cs+, не могут проходить сквозь канал дикого типа (Palmer L.G., 1990). Предполагается, что катионы проходящие через канал полностью лишены гидратной оболочки или связаны не более чем с одной молекулой воды в районе селективного фильтра (Doyle D.A. et al., 1998). Исходя из радиуса катионов K+ и Na+ было высказано предположение, что диаметр селективного фильтра у ENaC дикого типа составляет 1,9−2,7 Е. На основании исследования селективности каналов с мутациями G587 и S589 в б-субъединице, G529 в в-субъединице (положение гомологично бG587) и S542 в г-субъединице (положение гомологично бS589), предполагается, что данный участок непосредственно является селективным фильтром (Snyder P.M. et al., 1999; Kellenberger S. et al., 2001) (Рис. 10).
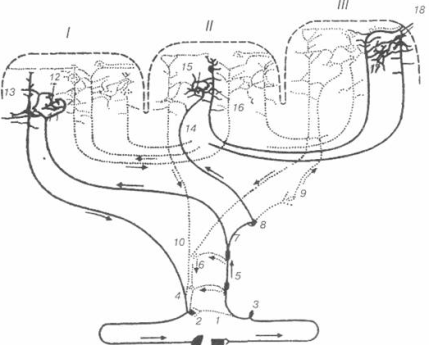
Рис. 10. Предсказанная структура поры и области короны ENaC. Предсказанные порообразующие трансмембранные домены гетеротримерного ENaC параллельно плазматической мембране (А). Консервативный Trp52 на внутриклеточный N-конце TM1 в-hENaC изображен с голубой боковой цепью. Консервативный Tyr (358 и 391) в предполагаемых петлях связи ви б-ENaC, которые находятся чуть выше поры, изображены с синими и желтыми боковыми цепями, соответственно. Изображение поры, перпендикулярно плазматической мембране с внеклеточной (В) и внутриклеточной © стороны, изображены консервативный Trp на N-конце TM1 и консервативный Ser (черные стрелки). На С-конце ТМ2, отмеченном для каждой субъединицы. Боковые цепи остатков в ТМ2 каждой субъединицы, образующей селективный фильтр, размещены на расстоянии, заполненном шариками. (модифицировано из Stockand et al., 2008)