Характеристика нового исходного материала галеги восточной, созданного методом полиплоидии

Перевод сортообразцов на тетраплоидный уровень проводили в ГНУ «Институт генетики и цитологии НАН Беларуси» путем обработки их семян раствором колхицина. Следует отметить, что семена белоцветкового сортообразца СЭГ-1 после всех обработок сильно разбухли и распадались, что не позволило использовать их для посева. У всех остальных сортообразцов из обработанных колхицином высеянных семян была… Читать ещё >
Характеристика нового исходного материала галеги восточной, созданного методом полиплоидии (реферат, курсовая, диплом, контрольная)
Характеристика нового исходного материала галеги восточной, созданного методом полиплоидии
Создание новых селекционных сортов методами массового и индивидуального отборов было серьезным этапом в селекции растений. Использование различных методов отбора в селекции галеги восточной позволяет выделить формы и биотипы лишь с теми признаками и свойствами, которые сформированы под действием естественного отбора, а селекционер, проводя искусственный отбор, лишь ускоряет этот процесс. Однако такие сорта в основном сохраняют признаки старых сортов. Для создания сортов с новыми признаками и свойствами необходимо использовать синтетические методы селекции, обеспечивающие получение генетической изменчивости в популяциях. Весьма эффективным методом для получения генетической изменчивости в популяциях многолетних бобовых трав является полиплоидия. В большинстве случаев полиплоидные формы отличаются положительными морфологическими, физиологическими, биохимическими и хозяйственно полезными признаками. По сравнению с диплоидными видами полиплоиды имеют более мощное развитие растений, характеризуются относительно увеличенными размерами клеток, различных органов и тканей и более крупными листьями, цветками, плодами и семенами [1, 2]. галега полиплоидия урожайность.
Анализ источников
Метод полиплоидии успешно используется в селекции люцерны посевной, клевера гибридного, клевера лугового [3]. Достаточно сказать, что за рубежом около половины, а в республике Беларусь более 40% сортов клевера лугового имеют тетраплоидный набор хромосом. Было установлено, что метод полиплоидии более эффективен на перекрестноопыляемых культурах с низким основным числом хромосом, у которых используются в производстве вегетативные, а не генеративные органы. Преимущества полиплоидных форм у таких культур заключаются в повышенной урожайности вегетативной массы, более высокой зимостойкости, устойчивости к болезням и повышенной кормовой ценности. Галега восточная относится к перекрестноопыляемым культурам с основным числом хромосом 8 (2n=16) и возделывается в производстве для получения вегетативной массы [4].
Поэтому целью наших исследований было создание нового исходного материала для селекции галеги восточной методом полиплоидии.
Методы исследования
Для получения полиплоидных форм галеги восточной использовали колхициновую методику, применяемую для получения полиплоидных форм рода Pisum [5]. С этой целью семена замачивали в водном растворе колхицина (0,02%). Подсчет хромосом галеги восточной проводился на временных препаратах по общепринятой методике.
Объектами для проведения полиплоидизации служили новые сортообразцы галеги восточной, созданные на кафедре селекции и генетики УО БГСХА: белоцветковые СЭГ-1 и СЭГ-1а, сиреневоцветковый СЭГ-2, голубоцветковый СЭГ-3, синецветковый СЭГ-4, фиолетовоцветковые с антоцианом СЭГ-5 и СЭГ-6. В качестве контроля, у которого не проводилась полиплоидизация, использовался сорт Нестерка.
Основная часть
Перевод сортообразцов на тетраплоидный уровень проводили в ГНУ «Институт генетики и цитологии НАН Беларуси» путем обработки их семян раствором колхицина. Следует отметить, что семена белоцветкового сортообразца СЭГ-1 после всех обработок сильно разбухли и распадались, что не позволило использовать их для посева. У всех остальных сортообразцов из обработанных колхицином высеянных семян была выращена рассада, количество которой варьировало в зависимости от образца в пределах от 6 у белоцветкового до 22 шт. у фиолетовоцветкового (табл. 1).
Таблица 1. Динамика появления всходов обработанных раствором колхицина семян и сохраняемость растений
Сортообразцы. | Количество взошедших и сохранившихся растений 15.06 в поле, шт. | ||||
11.05. | 25.05. | 12.06. | 15.06. | ||
СЭГ-1 — белоцветковый с изоляцией. | ; | ; | ; | ; | |
СЭГ-1а — белоцветковый. | |||||
СЭГ-2 — сиреневоцветковый. | |||||
СЭГ-3 — голубоцветковый. | |||||
СЭГ -4 — синецветковый. | |||||
СЭГ-5 — фиолетовоцветковый. | |||||
СЭГ-6 — фиолетовоцветковый. | |||||
Сорт Нестерка (стандарт). | |||||
Всего. | ; | ; | ; | ||
Перед уходом в зиму у всех сортообразцов сохранилось 82 растения, в том числе: СЭГ-1а — 4, СЭГ-2 — 9, СЭГ-3 — 4, СЭГ-4 — 17, СЭГ-5 — 22, СЭГ-6 — 7 и Нестерка — 19 растений. Из всех проанализированных на плоидность растений было выделено только 10 из СЭГ-4 — 3 растения (ТЕТ-1, ТЕТ-3, ТЕТ-8), из СЭГ-6 — 5 растений (ТЕТ-2, ТЕТ-4, ТЕТ-5, ТЕТ-6, ТЕТ-7), из СЭГ-1а и Нестерки по 1 растению (ТЕТ-9 и ТЕТ-10 соответственно), или 13,8%, которые имели тетраплоидный набор хромосом (2n=32).
В 2009 г. ранней весной тетраплоидные растения из ГНУ «Институт генетики и цитологии НАН Беларуси» были привезены в УО БГСХА и высажены на опытном поле кафедры селекции и генетики индивидуально с площадью питания 1,0Ч1,0 м. За каждым растением проводились фенологические наблюдения, описывались морфологические и хозяйственно полезные признаки, учитывалась семенная продуктивность.
Высаженные в поле 23 апреля 10 тетраплоидных растений галеги восточной прошли все фазы развития и к середине августа сформировали семена. Исключение составило растение ТЕТ-1, у которого бобы не сформировались совсем.
В первый год жизни у растений выявлены различия по высоте, варьирование которой находилось в пределах от 62 до 96 см (табл. 2).
Таблица 2. Характеристика хозяйственно полезных признаков полиплоидных растений галеги восточной
Название признаков. | ТЕТ-1. | ТЕТ-2. | ТЕТ-3. | ТЕТ-4. | ТЕТ-5. | ТЕТ-6. | ТЕТ-7. | ТЕТ-8. | ТЕТ-9. | ТЕТ-10. | ||
Количество стеблей. | ||||||||||||
Тип куста. | прямост. | полупр. | полупр. | полупр. | полупр. | полупр. | полупр. | полупр. | полупр. | прямост. | ||
Высота растений, см. | ||||||||||||
Количество междоузлий на 1 стебле. | ||||||||||||
Диаметр стебля, мм. | 5,2. | 5,9. | 6,4. | 5,5. | 3,8. | 5,5. | 5,1. | 6,2. | 5,5. | 5,4. | ||
Стебель: антоциановая окраска. | средняя. | сильная. | слабая. | средняя. | отсутств. | слабая. | отсутств. | средняя. | отсутств. | отсутств. | ||
Лист: | длина, см. | |||||||||||
Длина листочка, см. | 5,7. | 5,4. | 5,9. | 6,2. | 9,9. | 5,2. | 6,1. | 5,3. | 4,3. | 4,4. | ||
Ширина листочка, см. | 3,2. | 3,3. | 4,6. | 3,9. | 2,5. | 3,5. | 3,8. | 3,5. | 3,1. | 3,6. | ||
Окраска. | зеленая. | зеленая. | св. зеленая. | зеленая. | зеленая. | тем.зеленая. | св.зеленая. | св.зеленая. | зеленая. | зеленая. | ||
Силуэт. | открытый. | закрыт. | закрыт. | промеж. | промеж. | промеж. | открытый. | промеж. | промеж. | закрыт. | ||
Боб. | Форма. | изогнут. | изогнут. | изогнут. | прямой. | прямой. | изогнут. | прямой. | прямой. | прямой. | изогнут. | |
Антоциан. | ; | ; | ; | |||||||||
Окраска семян. | желто-зеленые. | оливк. | желтые. | желто-зеленые. | желтые. | оливк. | желто-зеленые. | грязно-желтые. | желтые. | желтые. | ||
Наиболее низкорослым (62 см) было растение ТЕТ-5, а наиболее высокорослым (96 см) — ТЕТ-4. Формирование куста и рост побегов у каждого растения проходили по-своему. Прямостоячий куст отмечен у растений ТЕТ-1 и ТЕТ-10, а у всех остальных он был полупрямостоячим. Количество стеблей в кусте в зависимости от растения варьировало в пределах от 5 до 12 штук.
Среднее число междоузлий на главном стебле составило по растениям 7−11 штук. Самый высокий показатель (11 междоузлий) отмечен у ТЕТ-1.
Проявились различия по толщине стебля, варьирующей в пределах от 5,1 мм (ТЕТ-7) до 6,4 мм (ТЕТ-3), и по окраске листьев, которая у ТЕТ-3, ТЕТ-7, ТЕТ-8 была светло-зеленой, у ТЕТ-1, ТЕТ-2, ТЕТ-4 и ТЕТ-5 — зеленой и у ТЕТ-6 — темно-зеленой с антоцианом. Длина листьев варьировала в зависимости от растения в пределах от 12 см (ТЕТ-5) до 19 см (ТЕТ-1, ТЕТ-3 и ТЕТ-7). Варьирование длины и ширины листочков в зависимости от растения находилось в пределах от 4,3 до 9,9 см и от 2,5 до 4,6 см соответственно. Окраска цветков у растений варьировала от светло-синей до фиолетовой. Окраска семян варьировала от желтой до оливковой, бобы были изогнутой или прямой формы с наличием или отсутствием антоциана [6].
На второй год (2010 г.) для увеличения учетной площади делянки кусты вегетативным способом рассадили. Погодные условия в 2010 г. сложились неблагоприятные, и из-за жаркой погоды растения были угнетенными, поэтому дальнейшие исследования проводились в 2011 г. В 2011 г. отрастание травостоя началось 18 апреля, через 42?46 дней в зависимости от формы наступила фаза бутонизации к 30 мая, а через 6?10 дней растения вступили в фазу цветения (табл. 3).
Таблица 3. Фенологические наблюдения за полиплоидными формами галеги восточной
Название формы. | Количество дней. | Вегетационный период, дней. | |||
отрастание-бутонизация. | бутонизация-цветение. | цветение-созревание. | |||
Нестерка st. | |||||
ТЕТ-1. | |||||
ТЕТ-2. | |||||
ТЕТ-3. | |||||
ТЕТ-4. | |||||
ТЕТ-5. | |||||
ТЕТ-6. | |||||
ТЕТ-7. | |||||
ТЕТ-8. | |||||
ТЕТ-9. | |||||
ТЕТ-10. | |||||
Созревание наступило в первой декаде августа, что составило 64−66 дней от фазы цветения. Таким образом, вегетационный период составил от 112 до 122 дней, что на 10−20 дней больше, чем у диплоидного сорта-стандарта Нестерка. Различие между формами по продолжительности вегетации составляет 10 дней, следовательно, их можно разделить две группы спелости? 112−116 и 117−122 дня.
В наших исследованиях был проведен учет динамики линейного роста растений. Было установлено, что более высокими темпами роста формы характеризовались в период бутонизации-начала цветения. Различия по среднесуточному приросту отмечены по фазам развития. Так, среднесуточный прирост растений составил: в фазе стеблевания? 1,0−2,8 см, в фазе ветвления? 2,8−5,6 см и в фазе бутонизации — 3,2−7,4 см (рисунок).
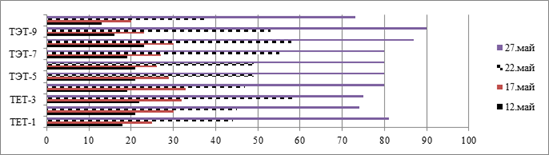
Рис. Динамика роста полиплоидных форм галеги восточной в 2011 г.
Ввиду того, что полученный материал весьма ценен, оценку урожайности зеленой массы не проводили, но оценку биологической урожайности зеленой массы, а также анализ облиственности и сухого вещества был проведен. Таким образом, при стеблестое 100 штук на 1 мІ урожайность зеленой массы составляет от 2,2 до 5,8 кг/мІ при содержании сухого вещества 14,3−23,4% (табл. 4).
Таблица 4. Биологическая урожайность зеленой массы, облиственность и содержание сухого вещества полиплоидных форм галеги восточной
Название формы. | Биологическая урожайность зеленой массы, кг/мІ. | Содержание сухого вещества, %. | Облиственность,%. | |
Нестерка st. | 4,4. | 14,5. | 45,5. | |
ТЕТ-1. | 5,8. | 17,9. | 62,6. | |
ТЕТ-2. | 2,8. | 19,7. | 61,8. | |
ТЕТ-3. | 5,8. | 22,8. | 49,1. | |
ТЕТ-4. | 3,7. | 20,7. | 54,1. | |
ТЕТ-5. | 2,2. | 23,4. | 60,5. | |
ТЕТ-6. | 4,3. | 20,4. | 57,0. | |
ТЕТ-7. | 3,0. | 19,5. | 50,7. | |
ТЕТ-8. | 4,3. | 21,2. | 54,1. | |
ТЕТ-9. | 5,1. | 19,9. | 54,5. | |
ТЕТ-10. | 3,5. | 14,3. | 52,2. | |
Наибольшей биологической урожайностью зеленой массы характеризовались ТЕТ-1 и ТЕТ-3 (5,8 кг/м), наименьшая урожайность отмечена у ТЕТ-5 (2,2 кг/мІ) и ТЕТ-2 (2,8 кг/мІ).
Все формы характеризовались высокой облиственностью, которая варьировала в пределах от 49,1 до 62,6%, причем формы ТЕТ-3, ТЕТ-9 и ТЕТ-10 имели необычный лист.
Важной характеристикой для нового сорта является семенная продуктивность. Учет семенной продуктивности проводился нами методом пробного снопа. При анализе снопа мы в своих исследованиях столкнулись с проблемой, с которой сталкивались и другие ученые.
Большинство экспериментально полученных аутополиплоидов характеризуется пониженной плодовитостью, что создает значительные трудности в селекции полиплоидов. В некоторых случаях это свойство, отрицательное в биологическом отношении, является желательным селекционным признаком. Частичная или полная бессемянность плодов у полиплоидов винограда и арбуза улучшает их качество. Уменьшение завязываемости семян при скрещивании диплоидов и тетраплоидов сахарной свеклы приводит к появлению у триплоидов ценного признака? резкого сдвига к односемянности клубочков, используемых для промышленных посевов. У культур, возделываемых не ради семян, частичная стерильность обычно не является препятствием к практическому использованию полиплоидов.
Однако в большинстве случаев, когда мы имеем дело с семенным воспроизводством, преодоление пониженной плодовитости является одной из главных трудностей в селекции полиплоидов.
У некоторых тетраплоидов отбор на повышение плодовитости быстро дает результаты. Так, у красного клевера плодовитость тетраплоидов составляла 32%; к третьему поколению ее удалось повысить до 53%. Путем отбора наиболее урожайных растений плодовитость была повышена до нормальной у тетраплоидов райграса многоукосного и пастбищного, овсяницы луговой и клевера розового [7, 8, 9].
В результате наших исследований у изучаемых полиплоидных форм галеги восточной к моменту созревания выявлено слабое варьирование по высоте растений и количеству междоузлий с коэффициентом вариации, равным 9,6 и 5,9% соответственно, или от 73 см у ТЕТ-10 до 102 см у ТЕТ-1 (табл. 5).
Таблица 5. Элементы структуры семенной продуктивности у полиплоидных форм галеги восточной
Название формы. | Высота стебля, см. | Кол-во междоузлий, шт. | Диаметр стебля, мм. | На один стебель. | Семян в бобе, шт. | Масса 1000 семян, г. | ||||
кистей. | бобов. | семян. | ||||||||
шт. | г. | |||||||||
Нестерка st. | 4,1. | 2,0. | 7,1. | |||||||
ТЕТ-1. | 4,8. | 1,5. | 4,3. | 7,2. | ||||||
ТЕТ-2. | 4,5. | 0,3. | 0,5. | 8,3. | ||||||
ТЕТ-3. | 4,4. | 0,03. | 0,7. | 5,6. | ||||||
ТЕТ-4. | 3,7. | 0,06. | 0,6. | 12,0. | ||||||
ТЕТ-5. | 5,3. | 0,5. | 0,9. | 8,2. | ||||||
ТЕТ-6. | 5,2. | 1,2. | 2,7. | 7,5. | ||||||
ТЕТ-7. | 6,2. | 0,2. | 0,7. | 9,4. | ||||||
ТЕТ-8. | 4,6. | 0,5. | 1,5. | 6,6. | ||||||
ТЕТ-9. | 4,5. | 0,2. | 0,9. | 11,0. | ||||||
ТЕТ-10. | 3,9. | 0−1. | 0,02. | 0,2. | 8,4. | |||||
Xmin. | 3,7. | 0,02. | 0,2. | 5,6. | ||||||
Xmax. | 6,2. | 1,5. | 4,3. | 12,0. | ||||||
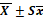 90±2,7. | 8,5±0,2. | 4,7±0,2. | 4,5±0,4. | 36,6±8,3. | 0,5±0,09. | 1,3±0,4. | 8,4±0,6. | |||
V%. | 9,6. | 5,9. | 15,4. | 28,9. | 72,4. | 66,5. | 96,9. | 22,6. | ||
Количество междоузлий на стебле составило 8−9 штук, варьирование диаметра стебля находилось в пределах от 3,7 до 6,2 мм (V=15,4%). Наибольшим диаметром характеризовалась форма ТЕТ-7 (6,2 мм). По всем остальным элементам семенной продуктивности отмечено сильное варьирование от 22,6 до 100%. В зависимости от полиплоидной формы на один стебель приходилось от 2 до 6 кистей (V=28,9%), на которых сформировалось от 2 до 73 бобов (V=72,4%). Наибольшее количество бобов отмечено у форм ТЕТ-6 (60 шт.), ТЕТ-5 (67 шт.) и ТЕТ-2 (73 шт.).
Наибольшее варьирование наблюдалось по количеству семян на одном стебле — от 1 до 208 штук, или от 0,02 до 1,5 г, которое достигало 100%. Наибольшее количество семян сформировали формы ТЕТ-6 (160 шт.) и ТЕТ-1 (208 шт.). Самая высокая стерильность отмечена у ТЕТ-10, на которой завязалось в среднем два боба на одном стебле, в которых сформировалось одно семя или вовсе ни одного.
Хорошую завязываемость семян имели формы ТЕТ-6 (2,7 шт.) и ТЕТ-1 (4,3 шт.).
Масса 1000 семян в зависимости от формы варьировала в пределах от 5,6 до 12,0 г, коэффициент вариации составил 22,6%. Самые крупные семена имели формы ТЕТ-9 и ТЕТ-4 с массой 1000 семян 11,0 и 12,0 г соответственно.
В результате проведенной оценки полиплоидных форм галеги восточной было установлено, что они различаются по продолжительности вегетационного периода и фенотипическим признакам. Наибольшими темпами роста характеризовались формы в период бутонизации-начала цветения, когда прирост составил 3,2−7,4 см за сутки.
Для повышения семенной продуктивности у полиплоидных форм необходимо вовлекать в скрещивания формы с высокими показателями элементов структуры урожайности семян на одном стебле: ТЕТ-2 и ТЕТ-7 с наибольшим числом кистей (6 шт.), ТЕТ-6, ТЕТ-5 и ТЕТ-2 — бобов (60−73 шт.), ТЕТ-6, ТЕТ-5 и ТЕТ-2 с наибольшим количеством завязавшихся семян (160−208 шт.).
Таким образом, созданный нами методом полиплоидии новый исходный материал характеризуется значительной изменчивостью не только качественных, но и количественных признаков и представляет практическую ценность для селекции новых сортов галеги восточной, более высокоурожайных по зеленой массе и семенам.
- 1. Таранухо, Г. И. Селекция и семенводство сльскохозяйственных культур: учеб. пособие / Г. И. Таранухо. — Минск: ИВЦ Минфина, 2009. — 420 с.
- 2. Порхунцова, О. А. Популятивность образцов галеги восточной и возможность их использования для отбора / О. А. Порхунцова // Сельское хозяйство — проблемы и перспективы: сб. науч. тр., Гродно, 14−15 марта 2005 г. / Гродн. гос. аграр. ун-т; под ред. В. К. Пестиса. — Гродно, 2005. — Т. 4, ч. 1. — С. 49−53.
- 3. Васильева, Т. В. Вредители нетрадиционных кормовых культур / Т. В. Васильева // Защита и карантин растений. — 2004. — № 3. — С. 56−57.
- 4. Бушуева, В. И. Галега восточная: монография / В. И. Бушуева, Г. И. Таранухо. — 2-е изд., доп. — Минск: Экоперспектива, 2009. — 193 с.
- 5. Дьячук, П. А. Экспериментальная полиплоидия в роде Pisum / П. А. Дьячук, И. С. Попова // Экспериментальная полиплоидия в селекции растений: сб. науч. тр. / А. Н. Лутков (отв. ред.) [и др.]. — Новосибирск: Наука, 1966. — С. 288−297.
- 6. Авраменко, М. Н. Создание исходного материала для селекции галеги восточной методом полиплоидии / М. Н. Авраменко, В. И. Бушуева // Молодежь и инновации — 2011: материалы Междунар. науч.-практ. конф., Горки, 25−27 мая 2011 г. / Белорус. гос. с.-х. акад.; редкол.: А. П. Курдеко [и др.]. — Горки, 2011. — Ч. 1. — С. 236−239.
- 7. Дубинин, Н. П. Теоретические вопросы и достижения при использовании полиплоидии в селекции растений / Н. П. Дубинин, В. К. Щербаков // Полиплоидия и селекция: сб. науч. тр., Москва, 14−18 янв. 1963 г. / Ин-т Цитологии Акад. наук СССР; отв. ред. П. М. Жуковский. — Л.: Наука, 1963. — С. 18−41.
- 8. Навалихина, Н. К. Генетические основы селекции тетераплоидного клевера красного / Н. К. Навалихина. — Киев: Наукова думка, 1973. — 132 с.
- 9. Навалихина, Н. К. Эксперементальное получение тетраплоидов у красного клевера Trifolium pretense L. / Н. К. Навалихина // Полиплоидия и селекция: сб. науч. тр., Москва, 14−18 января 1963 г./ Ин.-т Цитологии Акад. наук СССР; отв. ред. П. М. Жуковский. — Л.: Наука, 1963. — С. 277−284.