Флористическое районирование Астраханской области

После сартасской стадии Каспия произошло значительное понижение уровня моря. В этот период среднеазиатский или южноказахстанский гипотетический предок вышеперечисленных астрагалов по обширным литоралям активно заселил освободившуюся территорию западного, северного и восточного побережья Каспийского моря. Довольно четко вырисовывается обособление степного эндемичного для Ергеней и правобережья… Читать ещё >
Флористическое районирование Астраханской области (реферат, курсовая, диплом, контрольная)
Согласно схеме флористического районирования земного шара (Тахтаджян, 1978) и степей и пустынь юго-востока России (Сагалаев, 2001) территория Астраханской области входит в состав следующих фитохорионов: Голарктическое царство; Древнесредиземноморское подцарство; Ирано-Туранская область; Туранская провинция; Прикаспийский округ. Нами обоснованно выделены в пределах Прикаспийского округа следующие районы и подрайоны (рис. 2): 1. Волго-Ахтубинский район (1.1. Волгоградско-Ступинский подрайон; 1.2. Ахтубинско-Никольский подрайон; 1.3. Харабалинско-Енотаевский подрайон); 2. Бахтемиро-Кигачский район; 3. Приморско-дельтовый район; 4. Степной район; 5. Баскунчакский район; 6. Богдинский район; 7. Пустынный район; 8. Приморский район.
Выполненное нами выделение флористических районов согласуется в своих границах с районированием Республики Калмыкии (Бакташева, 2000) и является логичным его продолжением.
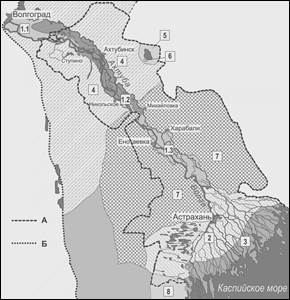
Рис. 2. Схематическая карта флористических районов и подрайонов Астраханской области и Прикаспийской провинции.
Условные обозначения: А — административная граница Астраханской области; Б — граница Прикаспийского округа проходящая по Ергеням (Сагалаев, 2001).
- 1. Волго-Ахтубинский район
- 1.1 Волгоградско-Ступинский подрайон
Южная граница подрайона определяется границей распространения дубрав с набором характерных для них видов (Rhamnus cathartica, Frangula alnus, Convallaria majalis, Scrophularia nodosa, Heracleum sibiricum, Vincetoxicum hirundinaria, Agrimonia asiatica, Acer tataricum, Viola himmetia, Alliaria petiolata, Viola accrescens, Hylotelephium triphyllum, Leucanthemum ircutianum, Cenolophium denudatum, Galium physocarpum, Sanguisorba officinalis, Pseudolysimachion longifolium и др.). Также южнее в пойме не отмечены и другие (Iris halophila, Potamogeton a…
После сартасской стадии Каспия произошло значительное понижение уровня моря. В этот период среднеазиатский или южноказахстанский гипотетический предок вышеперечисленных астрагалов по обширным литоралям активно заселил освободившуюся территорию западного, северного и восточного побережья Каспийского моря.
После образования наиболее молодой приморской группы бэровских бугров в дагестанскую стадию позднехвалынской трансгрессии гипотетический псаммофильный предок A. baerii, находившийся в зоне образования бэровских бугров, оказался изолированным на них (8 тыс. лет назад). Эндемичность A. baerii можно объяснить длительной «островной» изоляцией его гипотетического предка на самых южных (приморских) бэровских буграх. Длительная изоляция была вызвана тем, что более длительную «островную» историю имели самые южные по расположению бэровские бугры. Между ними располагались многочисленные протоки Волги (в период регрессий) и воды Каспийского моря (в период новокаспийской трансгрессии).
Довольно четко вырисовывается обособление степного эндемичного для Ергеней и правобережья Нижней Волги вида — Astragalus sytinii, как морфо-физиологического производного от A. lasiophyllus, которое сопровождалось заселением более мезофитных (чем среднеазиатские) степных экотопов юго-востока Русской равнины и Предкавказья.
Современные местонахождения A. sytinii полностью приуроченны к берегам бывшего Хвалынского моря, его максимальной и кумской стадиям позднехвалынской трансгрессии. Наибольшая вероятность проникновения родительского петрофильно-пустынного вида (A. lasiophyllus) на территорию Северо-Западного Прикаспия была во время енотаевской регрессии. В дальнейшем во время позднехвалынской трансгрессии (максимальной стадии) молодой таксон сохранился на берегах моря, в довольно прохладных и влажных условиях лесостепи. В промежуток времени между максимальной и кумской стадиями позднехвалынской трансгрессии ареал нового таксона, экологическим оптимумом которого являлся прохладный климат лесостепи, продвинулся на юг. Дальнейшему продвижению вида на юг, вслед за отступающим Хвалынским морем, воспрепятствовали, по-видимому, неподходящие эдафо-климатические условия.
С колебаниями уровня Каспийского моря связано образование галофильного вида Puccinellia choresmica. Вероятнее всего, что P. choresmica сформировался в результате «поглощения» путем гибридизации восточноприкаспийских популяций Puccinellia vitalii очень «активным» видом Puccinellia hauptiana.
Первоначально ареал P. vitalii включал территорию, простиравшуюся от Каспийского до Аральского моря. В результате трансгрессий Каспийского моря ареал Puccinellia vitalii оказался разорванным на две части (дезъюнктивный ареал). Одна часть ареала сохранилась в дельте р. Волги, вторая — в дельтах рек, впадающих в Аральское море. Именно в Аральской части ареала P. vitalii произошла гибридизация его с практически космополитным P. hauptiana. Изоляцией, способствующей сохранению P. vitalii в дельте Волги, по всей видимости, мы обязаны позднехвалынской трансгрессии. Возможным местом сохранения P. vitalii в период трансгрессий являлись бэровские бугры, где растения могли расти по нижней сырой части склонов, которые были берегами моря, т. е. в литоральных условиях.