Микротериокомплексы климатического оптимума голоцена как эталоны видового разнообразия при оценке трансформации рецентных биотопов Беларуси

На основании полученных результатов по благоприятности условий среды сделана попытка оценить трансформацию современных изученных биотопов по видовому разнообразию микромамма-лий. С этой целью автором введен новый показатель — коэффициент трансформации биотопов (Итб) по видовому разнообразию, который отражает величину, обратную индексу благоприятности (е) по отношению к соответствующим значениям… Читать ещё >
Микротериокомплексы климатического оптимума голоцена как эталоны видового разнообразия при оценке трансформации рецентных биотопов Беларуси (реферат, курсовая, диплом, контрольная)
Аннотация
В статье приведены результаты исследования приречных биотопов Национального парка, Полесского радиационно-экологического заповедника и ископаемой фауны микромаммалий атлантического периода голоцена.
Анализ имеющихся материалов показал, что современная и ископаемая микротериофауна имеют схожие структуру и видовой состав и представлены экологическими группами лесного и интразонального комплексов. Установлено, что микротериокомплексы современных приречных биотопов и сообществ атлантического периода голоцена имеют близкий видовой состав, что свидетельствует о серьезной трансформации приречных биотопов.
Оценка антропогенной трансформации территории, как в пределах отдельных регионов и отдельных компонентов природы, так и всей территории страны и ландшафтов в целом, чаще всего имеет относительный характер, так как при этом сравнивается с соответствующими показателями каких-либо эталонных территорий или ландшафтов. Однако вполне очевидно, что за исторический период времени на территории Беларуси, как и всего Европейского региона в целом, практически не осталось таких территорий, ландшафтов или отдельных биотопов, которые не были хотя бы частично в настоящее время или в прошлом преобразованы деятельностью человека. В связи с этим возникает необходимость дать абсолютную оценку трансформации ландшафтов или отдельных биотопов, сравнив их с соответствующими эталонными показателями до активного антропического воздействия на природную среду. заповедник биотоп фауна голоцен Изучение эволюции микротериокомплексов территории Беларуси [1−4] показало, что фауна микромаммалий, по сравнению с крупными животными, значительно меньше подвержена прямому антропогенному воздействию и испытывает на себе в основном косвенное влияние деятельности человека (через изменение растительных формаций и ландшафтов в целом). Поэтому практически до эпохи позднего неолита, т. е. второй половины среднего голоцена (суббореал) [5], изменение структуры и видового состава зооценозов определялось изменением климата и естественным ходом сукцессионной динамики ландшафтов, т. е. развитие микротериокомплексов до оптимума голоцена включительно (атлантический период голоцена) можно считать автохтонным, не зависящим от деятельности человека.
Светлохвойно-широколиственные и широколиственные леса, получившие широкое распространение в атлантическое время голоцена, в настоящее время на территории Беларуси характерны только для Полесья. Это позволяет сопоставлять приречные биотопы Полесского региона с эталонными атлантического времени по показателям видового разнообразия.
В ходе исследования изучалась рецентная (приречные биотопы Национального парка «Припятский» (НПП) и Полесского радиационно-экологического заповедника (ПРЭЗ) (всего 13 биотопов)) и ископаемая фауна микромаммалий атлантического периода голоцена (местонахождения Кирово, Пионерский (горизонт 1), Воронча, Заречье). Общее количество рецентных животных составило 3608 экземпляров, ископаемая фауна представлена 2096 определимыми остатками.
Анализ имеющихся материалов показывает, что современная и ископаемая микротериофауна имеют схожие структуру и видовой состав и представлены экологическими группами лесного и ин-тразонального (околоводного) комплексов (рис. 1). Однако для рецентной фауны НПП и ПРЭЗ по сравнению с оптимумом голоцена характерен более высокий процент видов открытых и лесолуговых биотопов (более 9%) за счет снижения удельного веса представителей широколиственных биотопов (с 31,5 до 2%). Данный факт уже сам по себе свидетельствует о серьезной трансформации приречных биотопов.
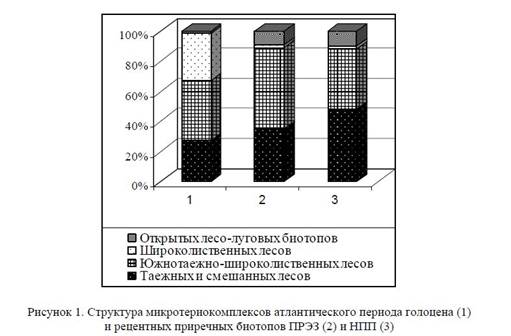
Структура микротериокомплексов атлантического периода голоцена (1) и рецентных приречных биотопов ПРЭЗ (2) и НПП (3).
Установлено, что микротериокомплексы современных приречных биотопов и сообществ атлантического периода голоцена имеют близкий видовой состав. Показатели сходства видового состава по индексу Серенсена рецентной и ископаемой микротериофауны хотя и несколько ниже рецентных, но довольно высоки и варьируют от 0,67 (ПРЭЗ) до 0,75 (НПП), что позволяет рассматривать эти фауны как сходные [6].
Наиболее очевидным показателем состояния любого биотопа является видовое разнообразие населяющих его организмов. При определении благоприятности условий среды по данным видового разнообразия использовались общепринятые показатели разнообразия: общее видовое разнообразие (Н) (индекс Шеннона), видовое богатство (d), выравненность (е), сходство видового состава сообществ (индекс Серенсена) и доминирование © (индекс Симпсона) [7].
О благоприятности условий экосистем наряду с количеством видов в сообществе (S) свидетельствуют кривые доминирования (рис. 2). Количественные характеристики доминирования отражает индекс доминирования (Симпсона) ©. Количественную оценку благоприятности среды отражает индекс выравненности (e), на основании которого можно определить благоприятность условий среды существования каждой фауны в градиенте между крайними положениями (при экстремальных условиях е = 0; при наиболее оптимальных е = 1) на основании следующей градации: 1−0,9 — условия оптимальные; 0,89−0,7 — мягкие; 0,69−0,5 — умеренные; 0,49−0,3 — суровые; 0,29−0,1 — близкие к экстремальным; 0,09−0,0 — экстремальные условия среды.
Значения благоприятности условий среды (e) атлантического времени рассматриваются как эталонные, по отношению к ним трансформация условий среды современных приречных биотопов по видовому разнообразию рассчитывалась как.
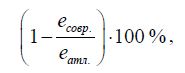
где есовр — показатель благоприятности условий среды современных приречных биотопов; еатл, — показатель благоприятности условий среды приречных биотопов климатического оптимума голоцена (АТ).
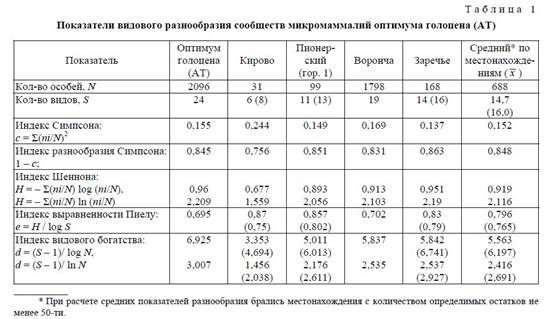
Анализ видового разнообразия современных и голоценовых сообществ микромаммалий показал, что для современных приречных биотопов Полесского региона характерно обеднение видового состава микротериокомплексов по отношению к оптимуму голоцена. Количество видов в рецентных биотопах колеблется от 7−8 до 11−12, а общее суммарное количество видов по всем выборкам региона составило 16 (18). Количество видов в ископаемых микротериокомплексах среднего голоцена существенно выше [8] и колеблется по местонахождениям от 12 до 19 видов, составляя в среднем 15, а общее суммарное количество видов из всех местонахождений этого периода — 24.
Кривая значимости видов атлантического периода максимально уплощена (рис. 2), доминантные виды отсутствуют, а среди содоминантов 3 вида: Clethrionomys glareolus Schreb. (23,1%), Sorex araneus L. (18,8%), в том числе и узкоспециализированный представитель широколиственных лесов Microtus subterraneus Sel.-Long. (23,7% остатков). Индекс доминирования © в сообществах среднего голоцена самый низкий за весь послеледниковый этап (средний по местонахождениям 0,175), а индексы разнообразия и выравненности максимально высокие (табл. 1), что указывает на очень благоприятные условия среды, которые по значению индекса выравненности (0,815) оцениваются как «мягкие».
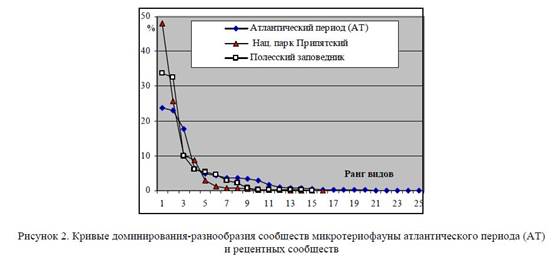
Кривые доминирования-разнообразия сообществ микротериофауны атлантического периода (АТ) и рецентных сообществ Начиная с эпохи позднего неолита (SB-SA) постепенно складывается система подсечного, а впоследствии и подсечно-огневого земледелия [6,7]. Неуклонное увеличение площади пашни, развитие земледелия и животноводства обусловили деградацию широколиственных лесов [2, 8−10].
В структуре микротериофауны приречных биотопов отмечается более высокий процент видов открытых лесо-луговых биотопов (более 9%); на фоне сокращения численности и обеднения видового состава лесного комплекса снижается до минимума удельный вес узкоспециализированных представителей широколиственных лесов (с 31,5 до 2%). В составе лесного комплекса появляются доминантные виды с высокими значениями доминирования (Clethrionomys glareolus Schreb.), удельный вес которых в структуре рецентных сообществ колеблется от 33 до 51% и более. Количество содоминантов при этом сокращается до 1−2 видов, которыми чаще всего являются Apodemus flavicollis Melch. и Microtus arvalis Pall., последняя, в зависимости от характера биотопов, может замещаться Sorex araneus L. или Microtus agrestis L. Кривые значимости видов (рис. 2) приобретают стремительное падение, а значения индекса доминирования почти в 2 раза превышают аналогичные показатели оптимума голоцена (средний показатель по биотопам составляет от 0,285 для ПРЭЗ до 0,334 для НПП) и сопоставимы с аналогичными показателями финальных этапов Валдайского оледенения и дриасовых стадиальных эпох позднеледниковья [4].
Здесь и далее в скобках указаны значения показателей разнообразия исходя из того, что при расчете индексов разнообразия современных сообществ микромаммалий некоторые виды (Brinaceus europaeus L,. Talpa europaea L.) не были учтены, так как не могли быть пойманы в давилки, хотя они и населяют биотопы речных долин. Вместе с тем ископаемые остатки этих видов довольно часто встречаются в голоценовых отложениях республики и учитывались при оценке видового разнообразия палеосообществ. Поэтому для большей объективности количество видов в современных сообществах мелких млекопитающих было увеличено исходя из характера биотопа.
Доминантов при этом сокращается до 1−2 видов, которыми чаще всего являются Apodemus flavicollis Melch. и Microtus arvalis Pall., последняя, в зависимости от характера биотопов, может замещаться Sorex araneus L. или Microtus agrestis L. Кривые значимости видов (рис. 2) приобретают стремительное падение, а значения индекса доминирования почти в 2 раза превышают аналогичные показатели оптимума голоцена (средний показатель по биотопам составляет от 0,285 для ПРЭЗ до 0,334 для НПП) и сопоставимы с аналогичными показателями финальных этапов Валдайского оледенения и дриасовых стадиальных эпох позднеледниковья [4].
Значения других показателей видового разнообразия для рецентных сообществ являются самыми низкими за всю послеледниковую историю их развития [11]. Особенно разителен контраст по значениям индекса видового богатства, который в рецентных сообществах сопоставим с аналогичными показателями дриасовых эпох позднего ледниковья, что свидетельствует об ухудшении благоприятности условий среды их существования. Низкие значения индексов разнообразия, в том числе и значение индекса выравненности Пиелу, которые в среднем составляют 0,635 (0,574) для биотопов НПП и 0,685 (0,66) — для ПРЭЗ, позволяют отнести оценку условий среды для мелких млекопитающих к градации «умеренные», однако по своему значению они более чем для других этапов голоцена смещены в сторону «суровых».
На основании полученных результатов по благоприятности условий среды сделана попытка оценить трансформацию современных изученных биотопов по видовому разнообразию микромамма-лий. С этой целью автором введен новый показатель — коэффициент трансформации биотопов (Итб) по видовому разнообразию, который отражает величину, обратную индексу благоприятности (е) по отношению к соответствующим значениям «эталонных биотопов», выраженную в процентах. В качестве «эталона» взяты показатели индекса благоприятности = выравненности (е) среды в оптимуме голоцена (АТ), среднее значение которых по местонахождениям этого этапа составляет 0,796 (0,765) (табл. 1).
Проведенная таким образом оценка трансформации биотопов по видовому разнообразию показала, что значения (Итб) для изученных рецентных биотопов варьируют от 6,2 (8,8) до 30,4 (34,1%), среднее значение показателя трансформации составило 17,1 (19,4%). В целом наиболее трансформированными по видовому разнообразию оказались биотопы пойм и надпойменных террас, представленные лесными широколиственными дубовыми и дубово-грабовыми формациями молодого возраста (табл. 2).
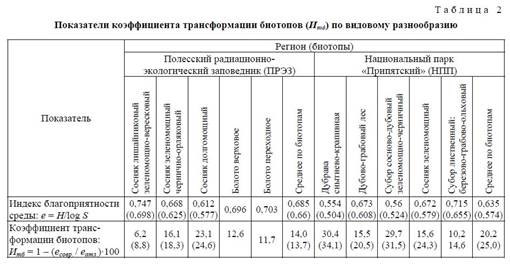
Таким образом, несмотря на то, что наибольшей продуктивностью обладают лесные формации широколиственных лесов, они же оказались и наиболее трансформированными по видовому разнообразию. Объясняется это тем, что, с одной стороны, коэффициент трансформации биотопов (Итб) рассчитывался на основе индекса выравненности видов в сообществе, а большинство микромаммалий указанных биотопов являются узковариабельными и довольно редкими видами с низкими показателями доминирования. Для отдельных из них (Microtus subterraneus Sel.-Long.) регион в настоящее время является северной границей распространения ареала. С другой стороны, на территории региона остается все меньше зрелых, наиболее продуктивных лесных широколиственных формаций, а молодые формации не обладают столь высокой продуктивностью и кормовой базой. В любом случае обе причины являются результатом антропогенной трансформации фитоценозов и, как следствие, трансформации структуры, состава и видового разнообразия микротериокомплексов.
- 1. Богдень И. И. История озер в СССР. — Таллинн: Ротапринт АН ЭССР, 1983. — Т. 10. — С. 36−38.
- 2. Гуман М. А. Антропогеновые изменения растительного покрова центральных районов Русской равнины в голоцене (по палинологическим данным): Автореф. дис. … канд. геогр. наук. — М., 1983. — 24 с.
- 3. Еловычева Я. К. Эволюция природной среды антропогена Беларуси. — Минск, 2001. — 292 с.
- 4. ИвановД.Л. Микротериофауна позднеледниковья-голоцена Беларуси. — Минск, 2008. — 215 с.
- 5. Иванов Д.Л.II Литосфера. — 2005. — № 2 (23). — С. 45−53.
- 6. Кабо В. Р. Первобытное общество и природа. — М.: Наука, 1981. — 152 с.
- 7. Калечиц Е. Г. Человек и среда обитания. Восточная Беларусь. Каменный век. — Минск, 2003. — 223 с.
- 8. Лийва А., Лозе И. Изотопно-геохимические исследования в Прибалтике и Белоруссии. — Таллинн, 1988. — С. 106
- 9. Motuzko А., Ivanov D. II Acta zool. cracov. — Krakow, — № 39 (1). — P. 381−386.
- 10. Одум Ю. Экология. — М., 1986. — Т. 2. — 376 с.
- 11. Савукинене Н. П., Сейбутис А. А. Палинология в континентальных и морских геологических исследованиях. — Рига, 1976. — С. 91−101.