Строение отделов головного мозга

Красные ядра, получая информацию от двигательной зоны коры головного мозга, подкорковых ядер и мозжечка о готовящемся движении и состоянии опорно-двигательного аппарата, регулируют тонус мускулатуры, подготавливая его уровень к намечающемуся произвольному движению. Черная субстанция связана с лежащими в основании полушарий переднего мозга базальными ганглиями — полосатым телом и бледным шаром… Читать ещё >
Строение отделов головного мозга (реферат, курсовая, диплом, контрольная)
Продолговатый мозг является непосредственным продолжением спинного мозга, у взрослого человека длина его около 25 мм. Он несколько сплющен в переднезаднем направлении и имеет форму усеченного конуса, суживающегося к спинному мозгу и расширяющегося к варолиеву мосту. По обеим сторонам от передней срединной щели продолговатого мозга расположены выпуклые белые тяжи — пирамиды, которые состоят из волокон еще общего здесь нисходящего кортикоспинального (пирамидного) тракта. Пирамиды суживаются книзу, около 2/3 их волокон постепенно переходят на противоположную сторону, образуя перекрест пирамид; спускаясь ниже, они образуют боковой кортикоспинальный путь. Меньшая часть волокон остается на той же стороне, переходя в передние канатики спинного мозга в виде переднего кортикосниналыюго пути (рис. 11.5).
Вдоль всего продолговатого мозга располагается ретикулярная формация, которая представлена переплетением нервных волокон и лежащими между ними нервными клетками. Ретикулярная формация связана восходящими и нисходящими волокнами с корой полушарий головного мозга, мозжечком и спинным мозгом, оказывая на кору полушарий и двигательные ядра спинного мозга активирующее воздействие.
Сбоку от пирамид выходит подъязычный нерв, корешки которого располагаются соответственно передним корешкам спинного мозга (см. Периферическая нервная система).
Боковые канатики занимают латеральные поверхности продолговатого мозга. Их вентральную (передненижнюю) часть составляют оливы, дорсальную (задневерхнюю) — нижние ножки мозжечка. Оливы имеют овальную форму и состоят из тел нейронов (ядра олив). Они функционально тесно связаны с мозжечком и ответственны за поддержание тела в вертикальном положении. Нижние ножки мозжечка — массивные волокнистые тяжи. Расходясь кверху в стороны, они ограничивают с боков нижний угол дна четвертого желудочка мозга — ромбовидную ямку. Все образования, расположенные между ромбовидной ямкой и пирамидами, относятся к покрышке.
Из боковых канатиков продолговатого мозга последовательно выходят корешки добавочного, блуждающего и языкоглоточного черепных нервов, располагающихся соответственно задним корешкам спинного мозга (см. Периферическая нервная система).
В нижней части на дорсальной (задней) поверхности продолговатого мозга тянется задняя серединная борозда, по бокам от которой заканчиваются утолщениями тонкий и клиновидный пучки задних канатиков спинного мозга. В утолщениях располагаются ядра этих пучков, отходящие.
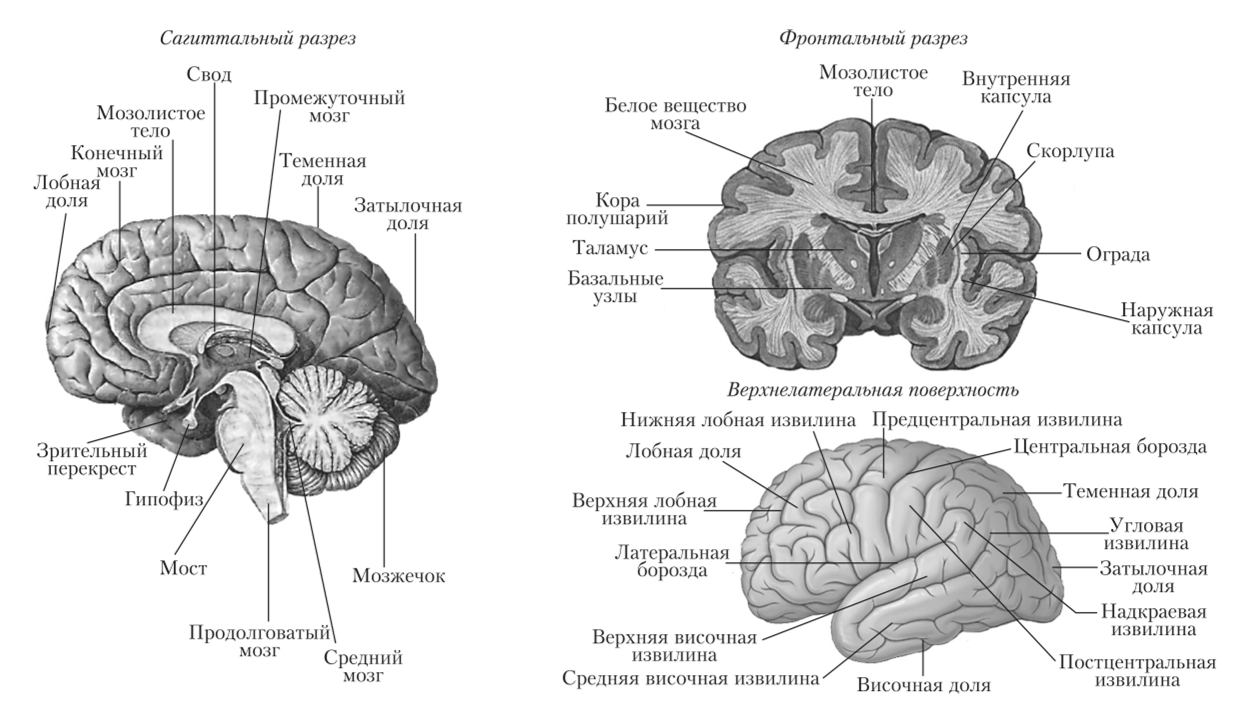
Рис. 11.4. Схема строения головного мозга.
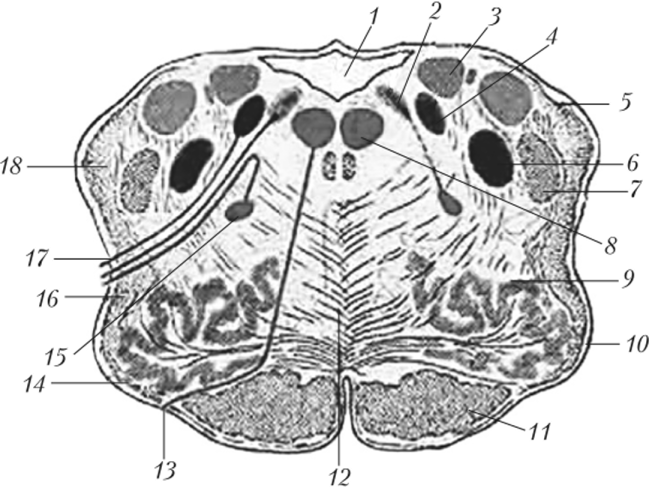
Рис. 11.5. Продолговатый мозг (поперечный разрез на уровне олив):
- 1 — четвертый желудочек; 2 — дорсальное ядро блуждающего нерва; 3 — ядро вестибулярного нерва; 4 — ядро одиночного пути; 5 — задний (дорсальный) спинномозжечковый путь; 6 — спинномозговое ядро тройничного нерва; 7 — спинномозговой путь тройничного нерва; 8 — ядро подъязычного нерва; 9 — оливное ядро;
- 10 — олива; 11 — корково-спинномозговой путь (пирамидный); 12 — медиальная петля; 13 — подъязычный нерв; 14 — передние наружные дуговые волокна;
- 15 — двойное ядро; 16 — спинно-таламический и спинно-покрышечный пути;
- 17 — блуждающий нерв; 18 — центральный (передний) спинно-мозжечковый путь
от них нервные волокна переходят па противоположную сторону в виде медиальной петли, направляясь затем к мосту, часть волокон входят в нижние ножки мозжечка. Через продолговатый мозг и нижние мозжечковые ножки проходят проприоцептивные пути мозжечкового направления — передний и задний спинно-мозжечковые.
Функции продолговатого мозга. Продолговатый мозг, так же как и спинной, выполняет две функции — рефлекторную и проводниковую. В продолговатом мозгу располагаются ядра следующих черепных нервов:
- — пара IX — языкоглоточный нерв; его ядро образовано тремя частями — двигательной, чувствительной и вегетативной. Двигательная часть участвует в иннервации мышц глотки и полости рта, чувствительная получает информацию от рецепторов вкуса задней трети языка; вегетативная иннервирует слюнные железы;
- — пара X — блуждающий нерв, имеет три ядра: вегетативное иннервирует гортань, пищевод, сердце, желудок, кишечник, пищеварительные железы; чувствительное получает информацию от рецепторов альвеол легких и других внутренних органов и двигательное (так называемое обоюдное) обеспечивает последовательность сокращения мышц глотки, гортани при глотании;
пара XI — добавочный нерв; его ядро частично расположено в продолговатом мозге; иннервирует грудиноключичнососцевидную и трапециевидную мышцы;
— пара XII — подъязычный нерв — двигательный нерв языка, его ядро большей частью расположено в продолговатом мозге.
Продолговатый мозг, гак же как и спинной, имеет чувствительную и двигательную связь с периферией. По чувствительным волокнам он получает импульсы от рецепторов кожи головы, слизистых оболочек глаз, носа, рта, от органа слуха, вестибулярного аппарата (органа равновесия), от рецепторов гортани, трахеи, легких, а также от интерорецепторов сердечно-сосудистой системы и системы пищеварения.
Через продолговатый мозг осуществляются многие простые и сложнейшие рефлексы, охватывающие многие жизнеобеспечивающие системы органов:
- — защитные рефлексы: кашель, чиханье, мигание, слезоотделение, рвота;
- — пищевые рефлексы: сосание, глотание, секреция пищеварительных желез;
- — сердечно-сосудистые рефлексы, регулирующие деятельность сердца и кровеносных сосудов;
- — рефлекторные центры дыхания: центр вдоха — инснираторный и центр выдоха — экспираторный, обеспечивающие автоматическую вентиляцию легких;
- — вестибулярные центры, обеспечивающие удержание позы тела вопреки земному притяжению.
Особое значение этого отдела центральной нервной системы определяется тем, что в продолговатом мозге находятся важнейшие центры жизнеобеспечения (дыхательный, сердечно-сосудистый и др.), поэтому не только удаление, но и даже повреждение продолговатого мозга заканчивается смертью.
Помимо рефлекторного продолговатый мозг выполняет проводниковую функцию. Через продолговатый мозг проходят проводящие пути, соединяющие двусторонней связью кору, промежуточный, средний мозг, мозжечок и спинной мозг.
Варолиев мост имеет вид поперечного валика, расположенного между средним мозгом сверху и продолговатым мозгом снизу. Дорсальная поверхность моста участвует в образовании ромбовидной ямки — дна IV мозгового желудочка. Вверху мост резко отграничен от ножек мозга. По бокам он суживается и переходит в средние мозжечковые ножки, уходящие в полушария мозжечка. Границей между средними мозжечковыми ножками и мостом является место выхода корешков тройничного нерва.
Мост отделяется от пирамид продолговатого мозга глубокой поперечной бороздой, из средней части которой выходят корешки правого и левого отводящих нервов (VI пара), а из латеральной (боковой) — корешки лицевого (VII пара) и преддверно-улиткового (VIII пара) нервов. Большая часть массы моста представляет собой белое вещество, т. е. скопления нервных волокон, образующих проводящие пути и черепные нервы.
Функции варолиева моста. Варолиев мост выполняет двигательные, сенсорные, интегративные и проводниковые функции. Важные функции моста связаны с наличием в нем ядер черепных нервов.
V пара — тройничный нерв (смешанный). Двигательное ядро нерва иннервирует жевательные мышцы, мышцы небной занавески и мышцы, напрягающие барабанную перепонку. Чувствительное ядро получает афферентные аксоны от рецепторов кожи лица, слизистой оболочки носа, зубов, 2/3 языка, надкостницы костей черепа, конъюнктивы глазного яблока.
VI пара — отводящий нерв (двигательный), иннервирует прямую наружную мышцу, отводящую глазное яблоко кнаружи.
VII пара — лицевой нерв (смешанный), иннервирует мимические мышцы лица, подъязычную и подчелюстную слюнные железы, передает информацию от вкусовых рецепторов передней части языка.
VIII пара — преддверно-улитковый (чувствительный) нерв. Улитковая часть этого нерва заканчивается в мозге в улитковых ядрах; преддверная — в треугольном ядре, ядре Дейтерса, ядре Бехтерева. Здесь происходит первичный анализ вестибулярных раздражений, их силы и направленности.
Через мост проходят все восходящие и нисходящие пути, связывающие мост с мозжечком, спинным мозгом, корой больших полушарий и другими структурами центральной нервной системы. По мостомозжечковым проводящим путям через мост осуществляется контролирующее влияние коры полушарий головного мозга на мозжечок. Кроме того, в мосте располагаются центры, регулирующие активность центров вдоха и выдоха, расположенных в продолговатом мозгу.
Мозжечок, или «малый мозг», располагается сзади от моста и продолговатого мозга. Он состоит из средней, непарной, филогенетически старой части — червя — и парных полушарий, свойственных лишь млекопитающим. Полушария мозжечка развиваются параллельно с корой больших полушарий и достигают у человека значительных размеров. Червь с нижней стороны расположен глубоко между полушариями; верхняя его поверхность переходит в полушария постепенно (рис. 11.6).
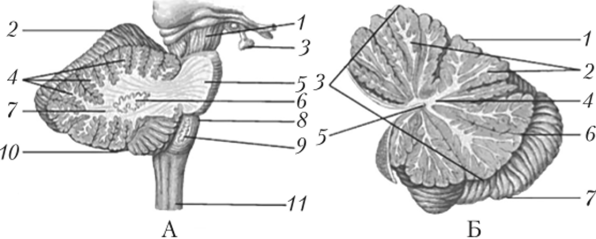
Рис. 11.6. Строение мозжечка (А — вид сбоку, Б — вертикальный разрез):
А: 1 — ножка мозга; 2 — верхняя поверхность полушария мозжечка; 3 — гипофиз; 4 — белые пластинки; 5 — мост; 6 — зубчатое ядро; 7 — белое вещество; 8 — продолговатый мозг; 9 — ядро оливы; 10 — нижняя поверхность полушария мозжечка; 11 — спинной мозг.
Б: 1 — верхняя поверхность полушария мозжечка; 2 — белые пластинки;
- 3 — червь; 4 — белое вещество; 5 — шатер; 6 — горизонтальная щель;
- 7 — нижняя поверхность полушария мозжечка
В целом мозжечок имеет обширные эфферентные связи со всеми двигательными системами стволовой части мозга: кортикоспинальной, рубросиинальной, ретикулоспинальной и вестибулоспинальной. Не менее разнообразными являются и афферентные входы мозжечка.
Вся поверхность мозжечка разделяется глубокими бороздами на доли. В свою очередь, каждая доля параллельными бороздками разделяется на извилины; группы извилин формируют дольки мозжечка. Полушария и червь мозжечка состоят из лежащего на периферии серого вещества — коры — и расположенного глубже белого вещества, в котором заложены скопления нервных клеток, образующие ядра мозжечка — ядра шатра, шаровидные, пробковидные и зубчатые.
Кора мозжечка имеет специфическое, нигде в ЦНС не повторяющееся строение. Все клетки коры мозжечка являются тормозящими, за исключением зернистых клеток самого глубокого слоя, которые оказывают возбуждающее воздействие.
Деятельность нейрональной системы коры мозжечка сводится к торможению нижележащих ядер, что предотвращает длительную циркуляцию возбуждения по нейронным цепям. Любой возбуждающий импульс, приходя в кору мозжечка, превращается в торможение за время порядка 100 мс. Так происходит как бы автоматическое стирание предшествующей информации, которое позволяет коре мозжечка участвовать в регуляции быстрых движений.
Функционально мозжечок можно разделить на три части: архиоцеребеллум (древний мозжечок), палеоцеребеллум (старый мозжечок) и неоцеребеллум (новый мозжечок). Архиоцеребеллум является вестибулярным регулятором, его повреждения приводят к нарушению равновесия. Функция палеоцеребеллума — взаимная координация позы и целенаправленного движения, а также коррекция выполнения относительно медленных движений по механизму обратной связи. При повреждении структур этой части мозжечка человеку трудно стоять и ходить, особенно в темноте, при отсутствии зрительной коррекции. Неоцеребеллум участвует в программировании сложных движений, выполнение которых идет без использования механизма обратных связей. В итоге возникает целенаправленное движение, выполняемое с большой скоростью, например игра на фортепиано. При нарушении структур неоцеребеллума нарушаются сложные последовательности движений, они становятся аритмичны и замедлены.
Мозжечок участвует в регуляции движений, делая их плавными, точными, соразмерными, обеспечивая соответствие между интенсивностью мышечного сокращения и задачей выполняемого движения. Мозжечок оказывает влияние также на ряд вегетативных функций, например, желудочно-кишечного тракта, на уровень кровяного давления, на состав крови.
Долгое время мозжечок считался структурой, ответственной исключительно за координацию движений. Сегодня признано его участие в процессах восприятия, когнитивной и речевой деятельности.
Средний мозг расположен над мостом и представлен ножками мозга и четверохолмием. Ножки мозга состоят из основания и покрышки, между которыми находится черная субстанция, содержащая сильно пигментированные клетки. В покрышке мозга располагаются ядра блокового (IV пара) и глазодвигательного (III пара) нервов. Полость среднего мозга представлена узким каналом — сильвиевым водопроводом, который соединяет III и IV мозговые желудочки. Длина среднего мозга у взрослого человека около.
2 см, вес — 26 г. В процессе эмбрионального развития средний мозг формируется из среднего мозгового пузыря, боковые выпячивания которого перемещаются вперед и образуют сетчатку глаза, структурно и функционально представляющая собой вынесенный на периферию нервный центр среднего мозга.
Наиболее крупными ядрами среднего мозга являются красные ядра, черная субстанция, ядра черепных (глазодвигательного и блокового) нервов и ядра ретикулярной формации. Через средний мозг проходят восходящие пути к таламусу, большим полушариям и мозжечку и нисходящие пути к продолговатому и спинному мозгу.
Средний мозг выполняет проводниковую, двигательную и рефлекторную функции.
Проводниковая функция среднего мозга заключается в том, что через него проходят все восходящие пути к вышележащим отделам: таламусу (медиальная петля, спиноталамический путь), большому мозгу и мозжечку. Нисходящие пути идут через средний мозг к продолговатому и спинному мозгу. Это пирамидный путь, корково-мостовые волокна, руброретикулоспинальный путь.
Двигательная функция среднего мозга реализуется за счет ядер блокового нерва, ядер глазодвигательного нерва, красного ядра, черной субстанции.
Красные ядра, получая информацию от двигательной зоны коры головного мозга, подкорковых ядер и мозжечка о готовящемся движении и состоянии опорно-двигательного аппарата, регулируют тонус мускулатуры, подготавливая его уровень к намечающемуся произвольному движению. Черная субстанция связана с лежащими в основании полушарий переднего мозга базальными ганглиями — полосатым телом и бледным шаром — и регулирует акты жевания, глотания (их последовательность), обеспечивает тонкую регуляцию пластического тонуса мышц и точные движения пальцев кисти руки, например, при письме. Нейроны ядер глазодвигательного и блокового нервов регулируют движение глаза вверх, вниз, наружу, к носу и вниз к углу носа. Нейроны добавочного ядра глазодвигательного нерва (ядро Якубовича) регулируют просвет зрачка и кривизну хрусталика. Со средним мозгом связано также осуществление выпрямительных и статокинетических рефлексов. Выпрямительные рефлексы состоят из двух фаз: подъема головы и последующего подъема туловища. Первая фаза осуществляется вследствие рефлекторных влияний рецепторов вестибулярного аппарата и кожи, вторая — связана с проприорсцспторами мышц шеи и туловища. Статокинетические рефлексы направлены на возвращение тела в исходное положение при перемещении тела в пространстве, при вращении.
Функционально самостоятельными структурами среднего мозга являются бугры четверохолмия. Верхние из них участвуют в деятельности первичных подкорковых центров зрительного анализатора, нижние — слухового. В них происходит первичное переключение зрительной и слуховой информации. Основная функция бугров четверохолмия — организация реакции настораживаиия и так называемых старт-рефлексов на внезапные, еще не распознанные, зрительные (верхнее двухолмие) или звуковые.
(нижнее двухолмие) сигналы. Активация среднего мозга при действии настораживающих факторов через гипоталамус приводит к повышению тонуса мышц, учащению сокращений сердца; происходит подготовка к избеганию или к оборонительной реакции. Кроме того, при нарушении четверохолмного рефлекса человек не может быстро переключаться с одного вида движения на другое.
Промежуточный мозг располагается под мозолистым телом и сводом, срастаясь по бокам с полушариями головного мозга. К нему относятся: таламус (зрительные бугры), гипоталамус (подбугорная область), эпиталамус (надбугорная область) и метаталамус (забугорная область) (рис. 11.7). Полостью промежуточного мозга является III желудочек мозга.
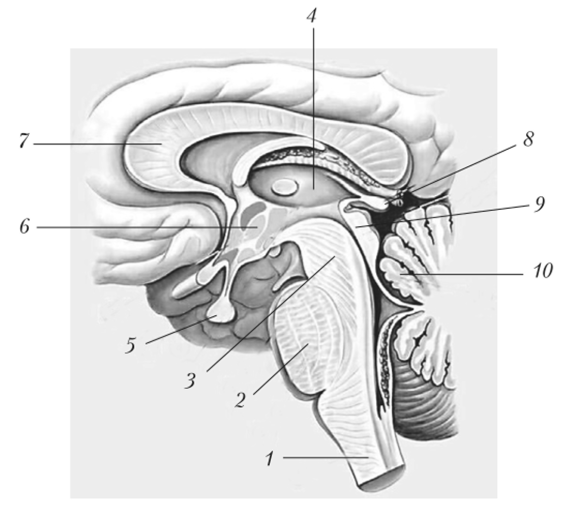
Рис. 11.7. Структуры, образующие ствол мозга (сагиттальный разрез):
- 1 — продолговатый мозг; 2 — мост; 3 — ножки мозга; 4 — таламус; 5 — гипофиз;
- 6' — проекция ядер подбугорной области; 7 — мозолистое тело; 8 — эпифиз;
- 9 — бугорки четверохолмия; 10 — мозжечок
Эпиталамус включает в себя железу внутренней секреции — эпифиз (шишковидное тело). В темноте она вырабатывает гормон мелатонин, который участвует в организации суточного ритма организма, влияет на регуляцию многих процессов, в частности на рост скелета и скорость полового созревания (см. Эндокринная система).
Метаталамус представлен наружными и срединными коленчатыми телами. Наружное коленчатое тело является подкорковым центром зрения, его нейроны по-разному реагируют на цветовые раздражения, включение, выключение света, т. е. могут выполнять детекторную функцию.
Срединное коленчатое тело — подкорковый, таламический центр слуха. Эфферентные пути от медиальных коленчатых тел идут в височную долю коры головного мозга, достигая там первичной слуховой зоны.
Таламус, или зрительный бугор, — парный орган яйцевидной формы, передняя часть которого заострена (передний бугорок), а задняя расширенная часть (подушка) нависает над коленчатыми телами. Срединная поверхность таламуса обращена в полость III желудочка мозга.
Таламус называют «коллектором чувствительности», так как к нему сходятся афферентные (чувствительные) пути от всех рецепторов, кроме обонятельных. В ядрах таламуса происходит переключение информации, поступающей от различных видов рецепторов, на начинающиеся здесь таламокортикальные пути, обращенные к коре головного мозга.
Главной функцией таламуса является интеграция (объединение) всех видов чувствительности. Для анализа внешней среды недостаточно сигналов от отдельных рецепторов. В таламусе происходит сопоставление информации, получаемой по различным каналам, и оценка ее биологического значения. В зрительном бугре насчитывается около 40 пар ядер, которые подразделяются на специфические (на нейронах этих ядер заканчиваются восходящие афферентные пути), неспецифические (ядра ретикулярной формации) и ассоциативные.
Отдельные нейроны специфических ядер таламуса возбуждаются рецепторами только своего типа. От специфических ядер информация о характере сенсорных стимулов поступает в строго определенные участки III—IV слоев коры головного мозга (соматотопическая локализация). Нарушение функции специфических ядер приводит к выпадению конкретных видов чувствительности, так как ядра таламуса, как и кора головного мозга, имеют соматотопическую локализацию. К специфическим ядрам таламуса идут сигналы от рецепторов кожи, глаз, уха, мышечной системы. Сюда же поступают сигналы от интерорецепторов зон проекции блуждающего и чревного нервов, гипоталамуса.
Нейроны неспецифических ядер образуют свои связи по сетчатому типу. Их аксоны поднимаются в кору головного мозга и контактируют со всеми ее слоями, образуя не локальные, а диффузные связи. К неспецифическим ядрам поступают связи из ретикулярной формации ствола мозга, гипоталамуса, лимбической системы, базальных ганглиев, специфических ядер таламуса. Усиление активности неспецифических ядер вызывает снижение активности коры головного мозга (развитие сонного состояния).
Сложное строение таламуса, наличие в нем взаимосвязанных специфических, неспецифических и ассоциативных ядер позволяет ему организовывать такие двигательные реакции, как сосание, жевание, глотание, смех, обеспечивать связь вегетативных и двигательных актов.
Через ассоциативные ядра таламус связан со всеми двигательными ядрами подкорки — полосатым телом, бледным шаром, гипоталамусом и с ядрами среднего и продолговатого мозга. Таламус является центром организации и реализации инстинктов, влечений, эмоций. Возможность получать информацию о состоянии множества систем организма позволяет таламусу участвовать в регуляции и определении функционального состояния организма в целом.
Гипоталамус (подбугорье) — структура промежуточного мозга, входящая в лимбическую систему и организующая эмоциональные, поведенческие, гомеостатические реакции организма. Гипоталамус имеет большое число нервных связей с корой головного мозга, подкорковыми узлами, зрительным бугром, средним мозгом, мостом, продолговатым и спинным мозгом. Ядра гипоталамуса имеют мощное кровоснабжение, его капилляры легко проницаемы для высокомолекулярных белковых соединений, что объясняет высокую чувствительность гипоталамуса к гуморальным сдвигам.
У человека гипоталамус окончательно созревает к 13−14 годам, когда заканчивается формирование гипоталамо-гипофизарных нейросекреторных связей. За счет мощных афферентных связей с обонятельным мозгом, базальными ганглиями, таламусом, гиппокампом, корой головного мозга гипоталамус получает информацию о состоянии практически всех структур мозга. В то же время гипоталамус посылает информацию к таламусу, ретикулярной формации, вегетативным центрам ствола мозга и спинного мозга.
Нейроны гипоталамуса имеют особенности, которые и определяют специфику функций самого гипоталамуса. К ним относятся отсутствие гематоэнцефалического барьера между нейронами и кровью, высокая чувствительность нейронов гипоталамуса к составу омывающей их крови и способность к секреции гормонов и нейромедиаторов. Это позволяет гипоталамусу воздействовать на вегетативные функции организма гуморальным и нервным путями.
В целом гипоталамус выполняет регуляцию функций нервной и эндокринной систем, в нем располагаются центры гомеостаза, теплорегуляции, голода и насыщения, жажды и ее удовлетворения, полового поведения, страха, ярости. Особое место в функциях гипоталамуса занимает регуляция деятельности гипофиза. В гипоталамусе и гипофизе образуются нейрорегуляторные вещества — энкефалины, эндорфины, обладающие морфиноподобным действием и способствующие снижению стресса.
Нейроны ядер передней группы гипоталамуса продуцируют вазопрессин, или антидиуретический гормон (АДГ), окситоцин и другие гормоны, которые по аксонам попадают в заднюю долю гипофиза — нейрогипофиз. Нейроны ядер срединной группы гипоталамуса продуцируют так называемые рилизинг-факторы, стимулирующие (либерины) и ингибирующие (статины) активность передней доли гипофиза — аденогипофиз, в котором образуются соматотропный, тиреотронный и другие гормоны (см. Эндокринная система). Нейроны гипоталамуса также обладают функцией детектора гомеостаза: реагируют на изменения температуры крови, электролитного состава и осмотического давления плазмы, количества и состав гормонов крови. Гипоталамус принимает участие в осуществлении половой функции и половом созревании, в регуляции цикла «бодрствование — сон»: задние отделы гипоталамуса активизируют бодрствование, стимуляция передних вызывает сон, повреждение гипоталамуса может вызвать так называемый летаргический сон.
Конечный мозг является наиболее молодым в филогенетическом отношении. Он состоит из двух полушарий, каждое из которых представлено плащом, обонятельным мозгом и базальными или подкорковыми ганглиями (ядрами). Длина полушарий в среднем — 17 см, высота — 12 см. Полостыо конечного мозга являются боковые желудочки, находящиеся в каждом из полушарий. Полушария головного мозга отделены друг от друга продольной щелью головного мозга и соединяются при помощи мозолистого тела, передней и задней спаек и спайки свода. Мозолистое тело состоит из поперечных волокон, которые в латеральном направлении идут в полушария, образуя лучистость мозолистого тела.
Обонятельный мозг представлен обонятельными луковицами, обонятельным бугорком, прозрачной перегородкой и прилежащими областями коры (препериформной, периамигдалярной и диагональной). Это меньшая часть конечного мозга, он обеспечивает функцию первого органа чувств, появившегося у живых существ, — функцию обоняния и, кроме того, входит в состав лимбической системы. Повреждение структуры лимбической системы вызывает глубокое нарушение эмоций и памяти.
Базальные ганглии (ядра серого вещества) расположены в глубине больших полушарий. Они составляют примерно 3% их объема. Базальные ганглии образуют многочисленные связи как между структурами, входящими в их состав, так и другими отделами мозга (корой больших полушарий, таламусом, черной субстанцией, красным ядром, мозжечком, двигательными нейронами спинного мозга). К базальным ганглиям относятся сильно вытянутое в длину и изогнутое хвостатое ядро и заложенное в толще белого вещества чечевицеобразное ядро. Двумя белыми пластинками оно подразделяется на скорлупу и бледный шар. Вместе хвостатое ядро и скорлупа носят название полосатого тела, связаны анатомически и характеризуются чередованием белого и серого вещества (рис. 11.8).
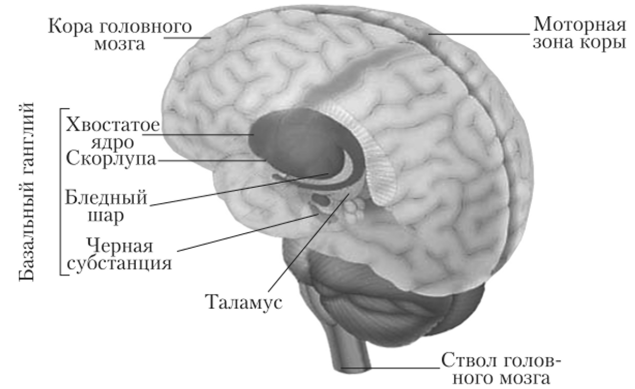
Рис. 11.8. Базальные ганглии.
Полосатое тело принимает участие в организации и регуляции движений и обеспечении перехода одного вида движения в другое. Стимуляция хвостатого ядра тормозит восприятие зрительной, слуховой и других видов сенсорной информации, угнетает активность коры, подкорки, безусловные рефлексы (пищевой, оборонительный и др.) и выработку условных рефлексов, приводит к наступлению сна. При поражении полосатого тела наблюдается выпадение памяти на события, предшествующие травме. Двустороннее повреждение полосатого тела побуждает к стремлению движения вперед, одностороннее — приводит к манежным движениям (ходьба по кругу). С нарушением функций полосатого тела связывают заболевание нервной системы — хорею (непроизвольные движения лицевых мышц, мышц рук и туловища). Скорлупа обеспечивает организацию пищевого поведения. При ее поражении наблюдаются трофические нарушения кожи, а ее раздражение вызывает слюноотделение и изменение дыхания. Функции бледного шара заключаются в провоцировании ориентировочной реакции, движения конечностей, пищевого поведения (жевание, глотание).
Плащ, или кора больших полушарий, — пластинка серого вещества, отделенная от полости желудочков белым веществом, которое содержит огромное количество нервных волокон, подразделяемых на три группы:
- 1. Пути, соединяющие различные отделы коры головного мозга внутри одного полушария, — ассоциативные пути. Выделяют короткие, или дугообразные, ассоциативные волокна, связывающие две лежащие рядом извилины, и длинные — протягивающиеся из одной доли в другую, оставаясь в пределах одного полушария.
- 2. Комиссуральные, или спаечные, волокна связывают кору обоих полушарий. Самой большой комиссурой головного мозга является мозолистое тело.
- 3. Проекционные пути связывают кору головного мозга с периферией. Существуют центробежные (эфферентные, двигательные) волокна, несущие нервные импульсы из коры на периферию, и центростремительные (афферентные, чувствительные) волокна, несущие импульсы с периферии в кору больших полушарий.
Кора больших полушарий является высшим отделом ЦНС. Она обеспечивает совершенную организацию поведения животных на основе врожденных и приобретенных в онтогенезе функций. Она делится на древнюю (archicortex), старую (paleocortex) и новую (neocortex). Древняя кора участвует в обеспечении обоняния и взаимодействия различных систем мозга. Старая кора включает поясную извилину, гиппокамп и участвует в реализации врожденных рефлексов и эмоционально-мотивационной сферы. Новая кора представлена основной частью коры больших полушарий головного мозга и осуществляет высший уровень координации работы мозга и формирования сложных форм поведения. Наибольшее развитие функций новой коры отмечается у человека, ее толщина во взрослом возрасте колеблется от 1,5 до 4,5 мм и максимальна в передней центральной извилине.
Морфологическое строение коры головного мозга. Кора состоит из многочисленных борозд и извилину благодаря которым поверхность коры значительно увеличивается. Они имеют индивидуальные различия не только у разных людей, но и в двух полушариях одного и того же человека. Глубокие, постоянные борозды деляг полушария на большие участки — доли} состоящие из долек и извилин. Долей всего шесть: лобная, теменная, височная, затылочная, краевая и островок (см. рис. 11.4).
Различают самые глубокие первичные борозды, которые делят полушария на доли. Боковая борозда (Сильвиева) отделяет лобную долю от височной, центральная борозда (Роландова) — лобную от теменной. Теменнозатылочная борозда располагается на медиальной поверхности полушария и разделяет теменную и затылочную доли, на верхнелатеральной поверхности явная граница между этими долями отсутствует. На медиальной поверхности полушария располагаются поясная, коллатеральная и обонятельные борозды. Поясная борозда идет параллельно мозолистому телу, отделяя лобную и теменную доли от поясной извилины. Коллатеральная борозда разграничивает на нижней поверхности полушария височную, краевую и затылочную доли. В передней части нижней поверхности полушария расположена обонятельная борозда с обонятельной луковицей, которая продолжается в обонятельный тракт.
В глубине боковой борозды располагается островковая доля. Она окружена с трех сторон круговой бороздой, ее поверхность изрезана бороздами и извилинами. Функционально эта доля связана с обонятельным мозгом.
Вторичные борозды менее глубокие, они делят доли на извилины и располагаются снаружи от одноименных извилин. Третичные (безымянные) борозды придают извилинам индивидуальную форму, увеличивают площадь их коры.
В лобной доле параллельно центральной борозде располагается предцентральная борозда. От нее в продольном направлении отходят верхняя и нижняя фронтальные борозды, которые делят долю па одну вертикальную и три горизонтальные извилины. Вертикальная извилина расположена между центральной и предцентральиой бороздами и называется предцентральной извилиной, в ней находится ядро двигательного анализатора. От пятого слоя коры этой извилины начинается корковый нисходящий путь. Горизонтальные извилины называются верхней, средней и нижней лобными извилинами. В средней извилине располагается центр письма — двигательный анализатор письменной речи, ядро которого окончательно формируется к 7 годам, а также центр сочетанного поворота головы и глаз в одну сторону. В нижней извилине локализован моторный центр речи (артикуляции) — центр Брокка, имеющий двустороннюю закладку в эмбриогенезе и развивающийся у правшей слева, а у левшей — справа. Ядро двигательного анализатора устной речи дифференцируется к 3 годам.
Теменная доля между центральной и постцентральной бороздами содержит постцентральную извилину, которая является центром осязания, болевой и температурной чувствительности. Перпендикулярно постцентральной извилине идет межтеменная борозда, разделяющая заднюю часть теменной доли на верхнюю и нижнюю теменные дольки. В верхней находится центр стереогнозиса (узнавания предметов на ощупь). В нижней теменной дольке видна надкраевая извилина, в которую упирается латеральная извилина. Надкраевая извилина является центром праксиса (целенаправленных действий, лежащих в основе формирования навыков различных видов деятельности). Ниже надкраевой лежит угловая извилина, где находится зрительный анализатор письменной речи (центр чтения), ядро которого формируется к 7-летнему возрасту. Два последних центра имеют двустороннюю закладку в эмбриогенезе, в дальнейшем развиваются у правшей слева, а у левшей — справа.
Височная доля имеет две продольные — верхнюю и нижнюю височные — борозды, которые делят се на три продольные извилины — верхнюю, среднюю и нижнюю. Все они параллельны латеральной борозде. В задней части верхней височной извилины выделяют сенсорный центр речи — центр Вернике. В среднем ее отделе располагается ядро слухового анализатора. У новорожденного оно подготовлено к восприятию различной звуковой стимуляции, но наиболее избирательно — к восприятию звуков человеческой речи. По мере развития речи корковый центр слуха быстро усложняется. В самой медиальной части располагается гиппокампальная извилина. Передний ее отдел представлен крючком, и здесь располагается центр обоняния и вкуса.
Затылочная доля имеет изменчивые и непостоянные борозды. На ее медиальной поверхности выделяется глубокая постоянная шпорная борозда, расположенная горизонтально и идущая от затылочного полюса до теменно-затылочной борозды. Между шпорной и затылочно-теменной бороздами находится треугольная извилина (клин) и язычная извилина — центр зрительного анализатора, ядро которого у новорожденного по своему клеточному составу сходно с ядром взрослых. Под влиянием внешних факторов происходит его дальнейшее усложнение.
Островок имеет форму треугольника, верхушка которого обращена вперед и вниз. Он находится в латеральной борозде и со всех сторон ограничен глубокой круговой бороздой, поверхность его покрыта короткими извилинами.
Краевая доля располагается на медиальной поверхности полушарий и включает в себя поясную и парагиппокампальную извилину. Поясная извилина начинается внизу бороздой мозолистого тела, а сверху — поясной бороздой, которая отделяет ее от лобной и теменной долей. Она активно участвует в формировании межполушарных связей и интегративной обработке информации путем передачи ее из одного полушария в другое. Парагиппокампальная извилина ограничивается сверху гиппокампальной бороздой, а снизу коллатеральной, отделяющей ее от височной доли. Передний конец парагиппокампальной извилины образует крючок, охватывая передний конец гиппокампальной борозды.
На внутренней поверхности коры выделяют ряд образований, которые относятся к лимбической системе. Эта система регулирует работу внутренних органов, эндокринных желез и обеспечивает эмоциональные реакции.
Лимбическая система (от лат. limbus — кромка, кайма) — область, расположенная между корой больших полушарий и продолговатым мозгом и как бы окаймляющая его (рис. 11.9). Состоит она из различных анатомически и функционально связанных образований головного мозга: ядер нервных клеток, располагающихся в передней области таламуса, гипоталамуса, миндалевидного ядра и гиппокампа, находящегося по соседству с миндалевидным ядром. К ней относят также обонятельную луковицу и поясную, гиппокампальную и зубчатую извилины. Они образуют кольцо над мозолистым телом.
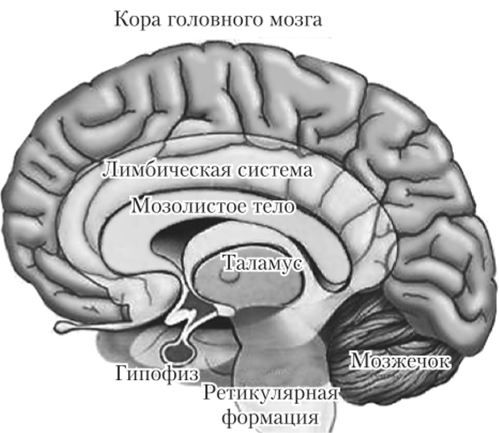
Рис. 11.9. Лимбическая система.
Основная функция лимбической системы — способность быстро приспосабливаться к изменениям внешней среды, быстро и адекватно реагировать на опасность. Главное место в этой приспособительной деятельности принадлежит эмоциям, биологический смысл которых как раз и заключается в быстрой оценке текущих потребностей организма и стимуляции соответствующего ответа на действие того или иного раздражителя. Кроме того, лимбическая система (в основном гиппокамп) принимает активное участие в сложнейших процессах, лежащих в основе памяти, преимущественно кратковременной.
Особенности строения коры головного мозга в онтогенезе. Взаимоотношения борозд и извилин с костями и швами черепа у новорожденного ребенка иные, чем у взрослого. Основные борозды (центральная, латеральная) выражены хорошо, но ветви основных борозд и мелкие извилины обозначены слабо. В дальнейшем в процессе развития коры борозды становятся глубже, а извилины между ними рельефнее. Соотношение борозд, извилин и швов черепа, характерное для взрослого человека, устанавливается у детей в 6—8 лет.
В течение первых месяцев жизни развитие коры идет очень быстрыми темпами. Большинство нейронов приобретают зрелую форму, интенсивно происходят процессы миелинизации нервных волокон, позволяющие реагировать на внешние раздражители более дифференцированно.
В процессе эволюции человека как биологического вида, а также в процессе онтогенеза — индивидуального развития каждого человека — происходит кортикализация функций, т. е. включение коры головного мозга в регуляцию функций нижележащих структур мозга. Это позволяет организовать более совершенную, учитывающую индивидуальный опыт, сохраненный в памяти, регуляцию функций организма. В дальнейшем, по мере автоматизации той или иной реакции, выполнение ее вновь передается подкорковым структурам с формированием автоматического реагирования.
Различные корковые зоны созревают неравномерно. Наиболее рано созревает соматосенсорная и двигательная кора, несколько позже — зрительная и слуховая. Особенно интенсивным в течение первого полугодия жизни является развитие зрительной коры, что влечет за собой развитие других зон мозга и их интеграцию. Созревание сенсорных и моторных зон в основном заканчивается к 3 годам. Значительно позже созревает ассоциативная кора: к 7 годам формируются ее основные связи, а окончательная дифференцировка, формирование нейронных ансамблей и связей с другими отделами мозга происходят к подростковому возраст}'. Наиболее поздно (ближе к 9 годам) созревают лобные области коры. Постепенность созревания структур коры больших полушарий определяет возрастные особенности высших нервных функций и поведенческих реакций детей различных возрастных групп.
Цитоархитектоника коры головного мозга. Общая площадь коры головного мозга человека около 2200 см2, число нейронов коры превышает 10 млрд. В составе коры имеются пирамидные, звездчатые, веретенообразные нейроны.
Пирамидные нейроны имеют разную величину, аксон пирамидного нейрона, как правило, проходит через белое вещество в другие зоны коры или в другие мозговые структуры.
Звездчатые клетки имеют короткие хорошо ветвящиеся дендриты и короткий аксон, обеспечивающий связи нейронов в пределах самой коры головного мозга.
Веретенообразные нейроны обеспечивают вертикальные или горизонтальные взаимосвязи нейронов разных слоев коры.
Кора головного мозга имеет преимущественно шестислойное строение (рис. 11.10).
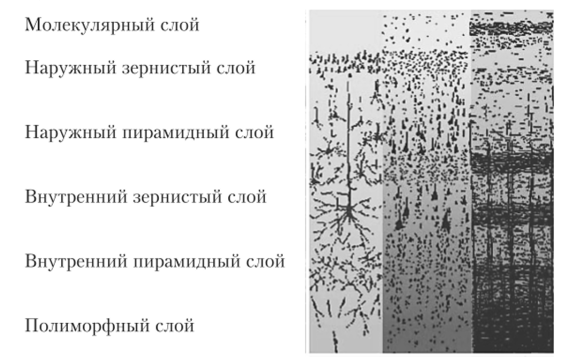
Рис. 11.10. Строение коры головного мозга.
Слой I — верхний молекулярный, представлен в основном ветвлениями восходящих дендритов пирамидных нейронов, среди которых расположены редкие горизонтальные клетки и клетки-зерна, сюда же приходят волокна неспецифических ядер таламуса, регулирующие через дендриты этого слоя уровень возбудимости коры головного мозга.
Слой II — наружный зернистый, состоит из звездчатых клеток, определяющих длительность циркулирования возбуждения в коре головного мозга, т. е. имеющих отношение к памяти.
Слой III — наружный пирамидный, формируется из пирамидных клеток малой величины и вместе со II слоем обеспечивает корко-корковые связи различных извилин мозга.
Слой IV — внутренний зернистый, содержит преимущественно звездчатые клетки. Здесь заканчиваются специфические таламокортикальные пути, т. е. пути, начинающиеся от рецепторов анализаторов.
Слой V — внутренний пирамидный (ганглиозный), слой крупных пирамид, которые являются выходными нейронами, аксоны их идут в ствол мозга и спинной мозг. В двигательной зоне в этом слое находятся гигантские пирамидные клетки, открытые Бецом (клетки Беца).
Слой VI — слой полиморфных клеток, большинство нейронов этого слоя образуют кортико-таламические пути.
Распределение нейронов по слоям в разных областях коры позволило выделить в мозге человека 53 цитоархитектонических поля (поля Бродмана), которые совершенствуются по мере развития коры головного мозга. У человека и высших млекопитающих различают, наряду с первичными, вторичные и третичные корковые поля, обеспечивающие ассоциацию функций данного анализатора с функциями других анализаторов.
Особенностью корковых полей является экранный принцип их функционирования, заключающийся в том, что рецептор проецирует свой сигнал не на один нейрон коры, а на поле нейронов, которое образуется их связями. В результате сигнал фокусируется не точка в точку, а на множестве разнообразных нейронов, что обеспечивает его полный анализ и возможность передачи в другие заинтересованные структуры. Так, одно волокно, приходящее в зрительную область коры, может активировать зону размером 0,1 мм. Это значит, что один аксон распределяет свое действие на более чем 5000 нейронов.
Функции отдельных зон новой коры определяются особенностями ее структурной организации, связями с другими структурами мозга, участием в восприятии, хранении и воспроизведении информации при организации и реализации поведения, регуляции функций сенсорных систем, внутренних органов.
Структурное различие участков коры головного мозга связано с различием их функций. В коре головного мозга выделяют сенсорные, моторные и ассоциативные области (рис. 11.11).
Корковые концы анализаторов имеют свою топографию — локальное расположение в определенных участках коры головного мозга. Они называются сенсорными областями коры головного мозга. Корковые концы анализаторов разных сенсорных систем перекрываются. Помимо этого, в каждой сенсорной системе коры имеются полисенсорные нейроны, которые реагируют не только на «свой» адекватный стимул, но и на сигналы других сенсорных систем. Эти механизмы лежат в основе формирования полимодальных связей, обеспечивающих сочетанную реакцию на различные раздражители.
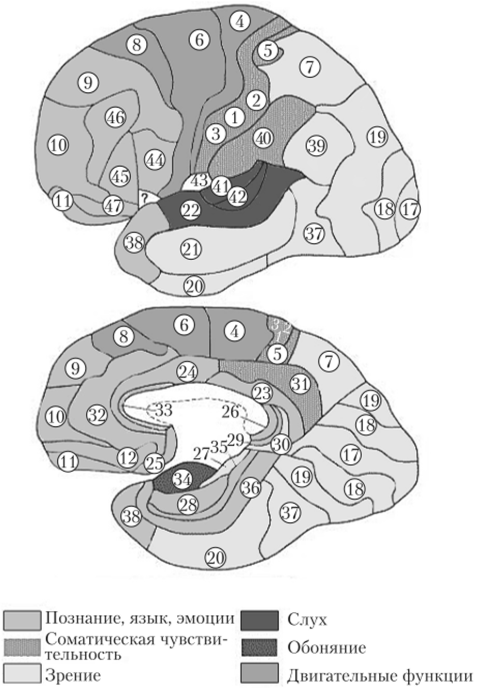
Рис. 11.11. Цитоархитектонические поля Бродмана.
Кожная рецептирующая система, таламокортикальные пути проецируются на заднюю центральную извилину. Здесь имеется строгое соматотопическое деление. На верхние отделы этой извилины проецируются рецептивные поля кожи нижних конечностей, на средние — туловища, на нижние отделы — руки, головы.
На заднюю центральную извилину в основном проецируется болевая и температурная чувствительность. В коре теменной доли (поля 5 и 7, см. рис. 11.11), где также оканчиваются проводящие пути чувствительности, осуществляется более сложный анализ: локализация раздражения, дискриминация, стереогноз. При повреждениях коры особенно сильно нарушаются функции дистальных отделов конечностей, особенно рук.
Зрительная система находится в затылочной доле мозга: поля 17, 18, 19. Центральный зрительный путь заканчивается в поле 17; он информирует о наличии и интенсивности зрительного сигнала. В полях 18 и 19 анализируются цвет, форма, размеры, качество предметов. Поражение поля 19 коры головного мозга приводит к тому, что больной видит, но не узнает предмет (зрительная агнозия, при этом утрачивается также цветовая память).
Слуховая система проецируется в поперечных височных извилинах (извилины Гешля), в глубине задних отделов латеральной (сильвиевой) борозды (поля 41, 42, 52). Именно здесь заканчиваются аксоны задних бугров четверохолмий и латеральных коленчатых тел.
Обонятельная система проецируется в области переднего конца гиппокампальной извилины (поле 34). Кора этой области имеет не шести-, а трехслойное строение. При раздражении отмечаются обонятельные галлюцинации, повреждение ее ведет к аносмии (потере обоняния).
Вкусовая система проецируется в гиппокампальной извилине по соседству с обонятельной областью коры (поле 43).
В передней центральной извилине расположены зоны, раздражение которых вызывает движение, они представлены по соматотопическому типу, но совершенно иначе: в верхних отделах извилины — нижние конечности, в нижних — верхние. Это моторные области коры головного мозга.
Спереди от передней центральной извилины лежат премоторные поля 6 и 8. Они организуют не изолированные, а комплексные, координированные, стереотипные движения. Эти поля также обеспечивают регуляцию тонуса гладкой мускулатуры, пластический тонус мышц через подкорковые структуры.
В реализации моторных функций принимают участие также вторая лобная извилина, затылочная, верхнетеменная области.
Двигательная область коры как никакая другая имеет большое количество связей с другими анализаторами, чем, видимо, и обусловлено наличие в ней значительного числа полисенсорных нейронов.
Все сенсорные проекционные зоны и моторные области коры занимают менее 20% поверхности коры головного мозга. Остальное — это ассоциативные области. Каждая ассоциативная область коры связана мощными связями с несколькими проекционными областями. В ассоциативных областях происходит интеграция разномодальной информации, позволяющей осуществлять осознание поступающей информации и сложные поведенческие акты. Ассоциативные области мозга у человека наиболее выражены в лобной, теменной и височной долях.
Каждая проекционная область коры окружена ассоциативными областями. Нейроны этих областей способны к восприятию разномодальной информации, обладают большими способностями к обучению. Полисенсорность нейронов ассоциативной области коры обеспечивает их участие в объединении поступающей информации, обеспечении взаимодействия сенсорных и моторных областей коры.
Так, в теменной ассоциативной области коры формируются субъективные представления об окружающем пространстве, о нашем теле. Это становится возможным благодаря сопоставлению соматосенсорной, проприоцептивной и зрительной информации. Лобные ассоциативные поля имеют связи с лимбическим отделом мозга и участвуют в организации программ действия при реализации сложных поведенческих актов с учетом их эмоциональной окраски.
Первой и наиболее характерной чертой ассоциативных областей коры является способность их нейронов воспринимать разномодальную информацию, причем сюда поступает не первичная, а уже обработанная информация с выделением биологической значимости сигнала. Это позволяет формировать программу целенаправленного поведенческого акта.
Вторая особенность ассоциативной области коры заключается в способности к пластическим перестройкам в зависимости от значимости поступающей информации.
Третья особенность ассоциативной области коры проявляется в длительном хранении следов сенсорных воздействий. Разрушение ассоциативной области приводит к выраженным нарушениям обучения, памяти.
Распределение функций по областям мозга не является абсолютным. Установлено, что практически все области мозга имеют полисенсорные нейроны, которые в определенной степени могут брать на себя функцию поврежденных модальноспецифических нейронов. Это позволяет компенсировать повреждение структур мозга в те периоды детства, когда поврежденная функция еще не закреплена жестко в структуре нервной ткани.
Важной особенностью коры головного мозга является ее способность длительно сохранять следы возбуждения. Это свойство придает коре исключительное значение в механизмах ассоциативной переработки и хранения информации, накопления знаний.
Межполугиариая асимметрия. Между правым и левым полушарием головного мозга существуют анатомические и функциональные различия. В результате нейропсихологических исследований было установлено, что полушария головного мозга различаются по функциональной специализации. В настоящее время считается доказанным, что с функциями левого и правого полушария у человека связаны два типа мышления — абстрактно-логическое и пространственно-образное и обозначаются они разными терминами:
- — вербальное и невербальное (поскольку абстрактно-логическое мышление в отличие от образного базируется на речевой деятельности);
- — аналитическое и синтетическое (поскольку с помощью логического мышления осуществляется анализ предметов и явлений, тогда как образное мышление обеспечивает цельность восприятия);
- — сукцессивное и симультанное (поскольку с помощью логического мышления осуществляется ряд последовательных операций, тогда как образное мышление обладает способностью к одномоментному восприятию и оценке объекта).
Известно также, что правополушарное мышление, создающее специфический пространственно-образный контекст, имеет решающее значение для творчества. Так, при органическом поражении левого полушария мозга у художников и музыкантов практически не страдают их артистические способности, а иногда даже повышается уровень эстетической выразительности творчества. Поражения правого полушария способны привести к полной утрате способности к творчеству. Вместе с тем все еще не выясненными остаются вопросы соотношения ведущей руки[1] и ведущего речевого полушария[2], связи межполушарной асимметрии с эмоциональной сферой и такими психическими познавательными процессами, как память и воображение.
Ведущим фактором в формировании межполушарной асимметрии считается генетическая предрасположенность, однако в некоторых случаях она может быть обусловлена прижизненными факторами, например, в результате легких повреждений мозга в процессе родов, приводящих к временному преобладанию функциональной активности того или иного полушария. Общепризнанным является положение о том, что межполушарная асимметрия проявляется нс только в предпочтении правой или левой руки, но и в целостной структурно-функциональной организации мозговой деятельности. В процессе онтогенеза межнолушарная асимметрия формируется в первые годы жизни и проявляется в первую очередь в выделении ребенком ведущей руки. Происходит это, как правило, в возрасте 2—3 лет, хотя в некоторых случаях несформированная латерализация (отсутствие в действиях отчетливого предпочтения той или иной руки) может сохраняться до 6—7 лет.
Следует отметить, что, несмотря на богатый фактический материал и активно ведущиеся исследования, единой теории, объясняющей все аспекты межполушарной функциональной асимметрии, до сих пор не существует. Однако не вызывает сомнения целесообразность функциональной асимметрии в сложной организации функций коры головного мозга, заключающаяся в повышении разнообразия адаптивных реакций и возможностей развития человеческих индивидуумов и всего человечества как биологического вида.