Описание обменных процессов в водной среде

В основу отечественных моделей водных (в частности, морских) экосистем легли работы основоположника российской математической экологии Алексея Андреевича Ляпунова по моделированию экосистемы пелагиали тропических вод океана. Модель была разработана А. А. Ляпуновым совместно с сотрудниками Института океанологии АН СССР для 44-го рейса исследовательского океанологического судна. Азот (Cn… Читать ещё >
Описание обменных процессов в водной среде (реферат, курсовая, диплом, контрольная)
В основу отечественных моделей водных (в частности, морских) экосистем легли работы основоположника российской математической экологии Алексея Андреевича Ляпунова по моделированию экосистемы пелагиали тропических вод океана. Модель была разработана А. А. Ляпуновым совместно с сотрудниками Института океанологии АН СССР для 44-го рейса исследовательского океанологического судна.
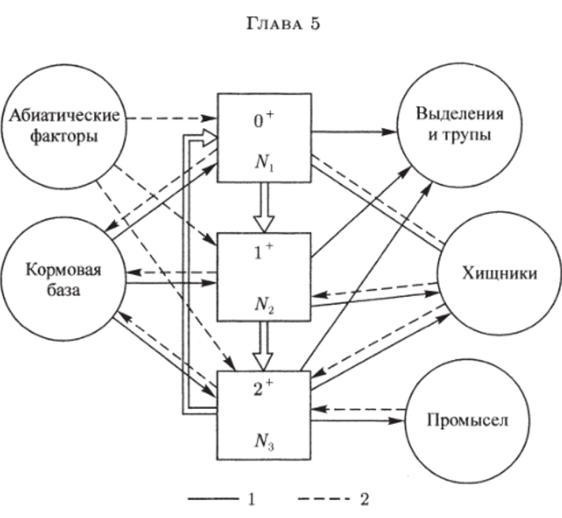
Рис. 5.1. Блок-схема популяции промысловой рыбы. Двойные стрелки — внутренние связи популяции: 1 — энергетические связи: 2 — информационные связи.
«Витязь», задача которого состояла в изучении биологической продуктивности пелагиали тропических вод океана в соответствии с международной биологической программой (Ляпунов, 1972; Виноградов, 1968).
Модель построена в предположении единообразия обменных процессов в водной среде и представляет собой систему дифференциальных уравнений. Основные представления о характере протекающих процессов легли в основу большого числа дальнейших моделей, описывающих конкретные водные экосистемы. На некоторых из них мы остановимся ниже.
В качестве компонент биогеоценоза пелагиали тропических вод были выделены следующие.
Солнечный свет — его объемная плотность L.
Усвояемый азот — его концентрация Сn.
Усвояемый фосфор — его концентрация С>.
Фитопланктон — его концентрация <�р.
Зоопланктон — его концентрация у?2.
Детрит — его концентрация ф.
Предполагалось, что другие биогенные вещества находятся в избытке и не играют лимитирующей роли. В некоторых случаях это предположение оказывается неправомерным.
Рассмотрим основные обменные процессы, протекающие в толще воды.
- • Поглощение света водой и взвешенными в ней телами.
- • Фотосинтез, осуществляемый фитопланктоном с использованием солнечной энергии и биогенных элементов. При этом интенсивность процесса биосинтеза определяется по принципу минимума (см. раздел 4.7).
- • Поедание фитопланктона зоопланктоном.
- • Каннибализм зоопланктона.
- • Поедание детрита зоопланктоном.
- • Размножение зоопланктона по схеме Вольтерра.
- • Отмирание зоопланктона и фитопланктона. Производство детрита зоопланктоном (трупы и экскременты).
- • Растворение детрита с образованием биогенных элементов.
- • Гравитационное оседание фитопланктона и детрит.
- • Собственное движение зоопланктона.
- • Диффузия всех составляющих, вызванная турбулентностью океанических вод.
В первом варианте изучаемой модели было выдвинуто предположение о том, что в горизонтальных плоскостях процесс развертывается единообразно. Движение океанических вод, кроме турбулентности, во внимание не принимаюсь.
Относительно каждой из компонент биогеоценоза были сделаны следующие допущения.
Солнечный свет (L) поглощается водой, фитопланктоном, зоопланктоном и детритом (коэффициенты соответственно а, аь а2, аз).
Свет принимает участие в фотосинтезе (коэффициент интенсивности фотосинтеза Л).
Азот (Cn) расходуется при биосинтезе фитопланктона. Предполагается, что суммарная интенсивность биосинтеза всегда пропорциональна интенсивности фотосинтеза (коэффициент расхода азота /in). Азот выделяется при разложении детрита (коэффициент x/n), а также подвергается диффузии, которая всецело вызвана турбулентностью (коэффициент диффузии Д).
Фосфор (Ср) участвует в тех же процессах, что и азот (коэффициенты /ip, ир, Д).
Фитопланктон (v?i) увеличивает свою биомассу (соответственно концентрацию) в процессе фотосинтеза (коэффициента). Он поедается зоопланктоном по «бимолекулярной» схеме (коэффициент (3) и подвергается гравитационному оседанию (коэффициент конвекции переноса вместе с водой иц, коэффициент турбулентной диффузии Д).
Зоопланктон (92) размножается по схеме Вольтерра. При этом он поедает' фитопланктон, сам себя (каннибализм), а также детрит (коэффициенты 71, 72, 73). Зоопланктон отмирает пропорционально своей концентрации в доле ?, подвергается диффузии с коэффициентом Д* > Д, так как он обладает собственными движениями, которые можно причислить к диффузии, кроме того, испытывает влияние турбулентной диффузии.
Наконец, зоопланктон совершает суточные миграции колебательного характера, в ночное время он поднимается, в дневное — опускается. Для разных слоев зоопланктона амплитуды миграции различны. В модели делается упрощающее предположение, что скорость конвекционного движения зоопланктона = const.
Детрит (ф) образуется при отмирании зсюпланктона (коэффициент v) и растворяется (коэффициент /х). Он также образуется из экскрементов зоопланктона в количестве, пропорциональном съеденной зоопланктоном пище, в виде фитопланктона, зоопланктона и детрита (коэффициенты ву #2, #з) — Детрит гравитационно оседает с коэффициентом о"з и подвергается турбулентной диффузии с коэффициентом Д.
Для определения суммарной интенсивности фотосинтеза (биосинтеза) применим подход, изложенный в гл. 4. В биосинтезе участвуют солнечная энергия, а также ряд веществ, например, N, Р, СО2, Са, S. Из эксперимента известно, что лимитирующую роль для процесса биосинтеза могут играть только азот, фосфор и световая энергия, так как остальные компоненты в тропических водах океана находятся в избытке. При этом нужно ввести коэффициенты, характеризующие использование лимитирующих веществ и световой энергии для синтеза единицы биомассы. Реальная интенсивность синтеза определяется лимитирующим фактором, т. е. той компонентой, которая обеспечивает наименьшую интенсивность синтеза.
Таким образом, существуют такие коэффициенты /, д^, <7р, что А = min(IL, 0nCn, ffpCp). Теперь можно записать систему уравнений:
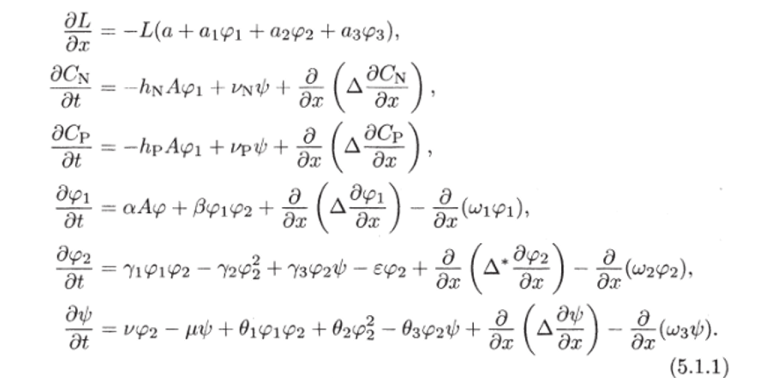
Знак минус перед членом 722 в уравнении для соответствует тому, что каннибализм меняет его массу за счет формирования детрита из зоопланктона.
Сформулируем краевые условия для этой системы уравнений. Решение ищем в полосе t > 0, < я < 0, где ~~ нижняя граница изучаемого слоя.
При х = 0 должна быть задана величина L (0, t). Кроме того, следует считать, что.
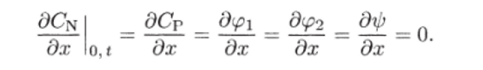
г-" gдСn дОр Otl%
При х = х должны быть заданы величинытг-^, -тг-, -тт—, -тг—, -=2-.
дх дх дх дх дх
При t = 0 должны быть заданы начальные концентрации Cn (x, 0), Cn (x, 0), Ср (х, 0), (pi (xy 0), ip2(x, 0), гр (х, 0).
Описанная выше модель сопоставлялась с экспериментальными результатами, полученными во время рейсов 1978 1986 гг. научно-исследовательского судна «Витязь». Можно отметить три основных пункта, в которых обнаружилось расхождение между гипотезами, лежащими в основе этой модели, и тем, что наблюдается в действительности.
- 1. Пренебрежение собственными движениями океанических вод следует считать недопустимым. Океанические воды состоят из ряда слоев, которые движутся поступательно в разных направлениях, что оказывает серьезное влияние на обменные процессы в системе.
- 2. Предположение о том, что в горизонтальных слоях процесс развертывается единообразно, далеко от действительности. Имеются «сгустки жизни» и области, в которых жизнь сильно разрежена.
- 3. Обменные процессы учтены в модели достаточно полно, однако необходимо считаться с тем, что минерализация детрита осуществляется бактериями. В таком случае нужно ввести в модель еще одну составляющую — бактерии. При этом агрегаты достаточно больших размеров, составленные из бактерий, сами могут служить нищей зоопланктону.
При конкретизации модели приходится учитывать известные из экспериментов и наблюдений сведения о параметрах системы и о виде функций взаимодействия компонентов биогеоценоза. Поскольку относительно входящих в уравнения (5.1.1) параметрах известно очень мало, для описания некоторых величин приходится использовать эмпирические формулы.
Например, известно, что скорость фотосинтеза на глубине 2 зависит от температуры воды Й, концентрации биогенов uz и биомассы фитопланктона. При этом др/dz —> 0, когда иг —? 0 и dp/dz —> const с возрастанием иг. Известно также, что скорость фотосинтеза как функция освещенности имеет один максимум при некотором оптимальном значении ?тах, убывая при увеличении и уменьшении освещенности от этого критического значения. Максимум фотосинтеза в тропических широтах в разные сезоны года располагается, как правило, на глубинах 10 30 м, в редких случаях (в открытых районах при повышенной прозрачности воды) он может наблюдаться на глубине ниже 30 м.
Анализируя наблюдающиеся в реальных условиях зависимости от интенсивности фотосинтеза от различных гидрологических, абиотических и биотических факторов, можно принять следующую приближенную формулу для описания фотосинтеза на глубине z (рис. 5.2):
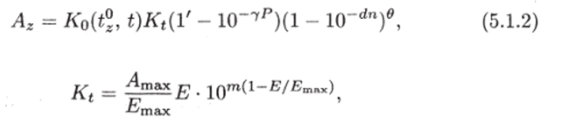
где.
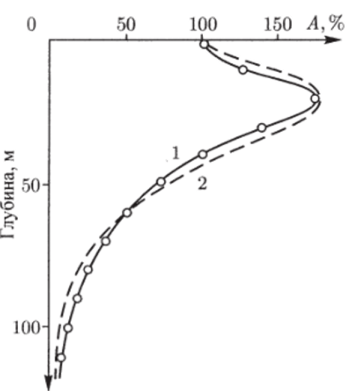
Рис. 5.2. Зависимость интенсивности фотосинтеза от вертикальной координаты: 1 — данные нолевых наблюдений; 2 — модельная кривая (Крапивин, 1978).
Kt — коэффициент пропорциональности; Ko (t^, t) — функция, характеризующая зависимость скорости фотосинтеза от температуры воды и времени; Лшах — ассимиляционное число в области максимального фотосинтеза (прирост на единицу веса фитоиланктонных организмов); га, 7, rf, в — константы.
Согласно многочис-зенным экспериментальным данным, ассимиляционное число тропического фитопланктона в области максимального фотосинтеза в среднем составляет около 11−12 мгС/ч. При этом световое насыщение в экваториальных районах Атлантического океана достигается при 9 кал/см2ч.
Таким образом, вместо величины А в формулах (5.1.2) для более точного соответствия приходится использовать величину фотосинтетической продукции фитопланктона, вычисленную в соответствии с системой уравнений (5.1.1).
Также пришлось конкретизировать функции рациона зоопланктона, причем весь зоопланктон разбить на несколько групп видов, образующих трофическую пирамиду. Гидрологические факторы также оказываются очень важными, в некоторых районах необходимо учитывать явление апвеллинга (вертикальных течений) или существование гермоклина.
Полная модель требует включения все больших подробностей и становится с трудом обозримой. Ясно, что степень детализации должна определяться как особенностями объекта, так и целью моделирования. Именно концептуальная схема, а не возможности компьютеров, определяют в настоящее время, как правило, ценность модели. Ниже мы остановимся на некоторых моделях водных систем различной степени сложности и тех теоретических и практических вопросах, которые можно решать с помощью динамических моделей.