Наземные беспозвоночные буферной зоны антропогенных полупустынных ландшафтов Северного Прибалхашья

За счет ортоптероидных насекомых живет большая групга хищных (скутигеры, тарантулы, пауки-скакуны, богомолы из рода Mantis, Bolivaria, Rivetina, мухи-ктыри и др.) и паразитических насекомых. Значительная часть энтомофагов — наездники. На стадии личинки они — паразиты личинок и куколок многих групп насекомых, в том числе сельскохозяйственных вредителей. Обычными хозяевами ихневмонид являются… Читать ещё >
Наземные беспозвоночные буферной зоны антропогенных полупустынных ландшафтов Северного Прибалхашья (реферат, курсовая, диплом, контрольная)
Беспозвоночные животные являются важной и мало изученной составляющей полупустынных экосистем Казахстана. Данные по энтомофауне, а также другим группам традиционно используют для биоиндикации антропогенного и техногенного загрязнения. Фауна беспозвоночных района Северного Прибалхашья изучалась в связи с отчуждением земель для восстановления деятельности промышленного комплекса. В качестве неизмененного природного ландшафта была выделена буферная зона. Исследование герпетобионтов на уровне крупных таксонов показало зависимость распределения беспозвоночных от экологического состояния биоценозов. Выполнен анализ динамической плотности и трофической структуры беспозвоночных.
Население наземных насекомых и других беспозвоночных полынно-солонцовых песчаных псаммофитных полупустынь Северного Прибалхашья изучено еще недостаточно. Информация о биоразнообразии групп отражена в таксономических статьях и монографиях различных специалистов Казахстана и других стран. Однако вопросы фаунистики, зоогеографии, экологии, фенологии, биологии этих групп в регионе еще полностью не выяснены ввиду отдаленности региона от научных зоологических центров. Возможно, многие виды беспозвоночных до сих пор не известны науке. Решение этих вопросов, в частности создание кадастра насекомых, могло бы способствовать оперативному выявлению, прогнозированию и предотвращению обеднения биологического разнообразия экосистем Северного Прибалхашья. Целью проведенных нами исследований являлось установление закономерностей распространения фауны наземных беспозвоночных в условиях полупустынных ландшафтов, в том числе антропогенно преобразованных.
Исследуемый район Северного Прибалхашья отличается большой засушливостью и континен-тальностью, в связи с чем воздействие стрессовых антропогенных факторов на флору и фауну проявляется в этом регионе с большей силой. Здесь расположены многочисленные предприятия горнодобывающей промышленности, для которых характерно интенсивное воздействие на окружающую природную среду, неизбежно вызывающее ее изменение. В процессе производства нарушается полностью или частично сложившееся экологическое равновесие в зонах размещения промышленных объектов (шахт, рудников, обогатительных фабрик). Эти изменения проявляются в отчуждении территорий для производства горных работ, истощении и загрязнении подземных и поверхностных вод, затоплении и заболачивании подработанных территорий, обезвоживании и засолении почв, загрязнении вредными веществами и химическими элементами атмосферного воздуха, изменении микроклимата. В связи с этим особую актуальность и значимость приобретают вопросы практического изучения фауны «культурного ландшафта», ее таксономического состава и направлений трансформации, которые происходят при широком освоении новых земель и их природных ресурсов.
Специфика влияния конкретного горнодобывающего предприятия на окружающую среду обусловлена геолого-геохимическими особенностями месторождений и применяемой техникой и технологией для его разработки. Распространение загрязняющих веществ в технологических цепях связано с технологией добычи и обогащения полезных ископаемых. Техногенные изменения окружающей среды при разработке месторождений полезных ископаемых, в особенности если она ведется длительное время, захватывают значительные территории, по площади несопоставимые с площадями горных отводов.
Таким образом, в совокупном проявлении большого комплекса техногенных процессов в районе горнодобывающих предприятий формируется техногенез горного профиля, в результате интенсивного воздействия которого происходит преобразование верхней части литосферы и окружающей среды в целом. При оценке качества природной среды и ее изменения в результате техногенного воздействия в качестве важных характеристик рассматривается состояние фауны почв, структура комплекса герпетобионтов и устойчивость его к техногенному воздействию, возможный уровень стабилизации процессов деградации фауны [1].
Материалы и методы исследования. Полевые работы были проведены в районе золотодобывающего месторождения «Пустынное» в Северном Прибалхашье в течение осени и весны 2011;2012 гг.
Территория месторождения находится в природной зоне боялычевых пустынь. Располагаясь на границе между степной и пустынной зонами, она совмещает в себе признаки тех и других в виде чередующихся злаковых (степных) и полынных (полупустынных) растительных сообществ, образующих пестрые сочетания [2].
Растительный покров исследованной территории характеризуется неоднородной пространственной структурой, довольно высоким биоразнообразием на видовом, популяционном, фитоценотиче-ском уровнях. Обычны белоземельно-полынно-боялычевые, тырсиково-белоземельно-полынные сообщества. Светло-каштановые почвы чередуются с сероземами. На большей части района почвы отличаются большой сухостью и засоленностью. В таких условиях развивается скудная растительность, а злаки играют подчиненную роль. Разреженный травостой низкорослый, часто встречаются оголенные участки, покрытые налетами солей [3].
В условиях резкого недостатка влаги даже малейшие, неуловимые для глаз неровности поверхности создают различия в водном режиме верхних горизонтов и, следовательно, в почвообразовательном процессе. Растения чутко реагируют на малейшие изменения среды. Этими причинами и объясняется пятнистость полупустыни в районе исследования. Растительный покров разреженный, на плакорных пространствах и повышениях он образован преимущественно полынями.
В понижениях встречаются боялыч, биюргун и терескен, а также льнянки, парнолистник. Каждой разновидности почв соответствует своя растительность. На светло-каштановых почвах растут типчак, белая полынь, ромашник. На лугово-каштановых — ковыль, типчак, на солонцах — полынь, прутняк, на солончаках развиваются солянки — сарзан, солерос. Во всех этих растительных сообществах заметную роль в травостое играют мятлик луковичный и полынок.
Нами проводились визуальные наблюдения, маршрутные учеты беспозвоночных, а также ручной сбор насекомых и учет ловушками на экспериментальных площадках буферной зоны, характеризующейся минимальной степенью техногенного воздействия.
Буферная зона представляет собой холмистую территорию с растительностью злаково-кустар-никового типа и проективным покрытием 80−90%. Растительность распространена довольно равномерно, изредка встречаются каменистые участки и островки, лишенные растений, с явными признаками сильного засоления. На территории буферной зоны доминантными видами являются боялыч, лебеда седая, тасбиюргун, курчавка, ковыль, очень обилен ревень татарский.
При выполнении визуальных наблюдений, маршрутных учетов беспозвоночных, а также учетов ловушками на экспериментальных площадках использовались традиционные почвенно-зоологиче-ские методики [4]. Всего за время проведения исследований было поставлено более 120 ловушек. Обработано и определено около 400 экземпляров беспозвоночных.
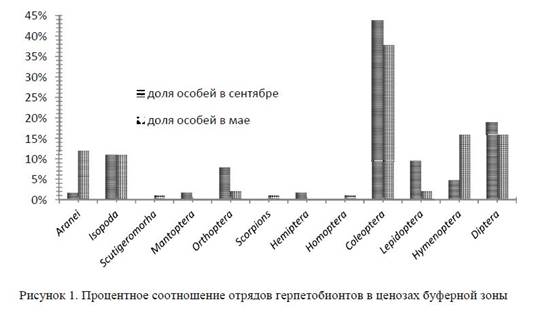
Результаты и их обсуждение. В результате исследований в биоценозах буферной зоны в сентябре нами были обнаружены представители 9 отрядов: Aranei, Isopoda, Mantoptera, Orthoptera, Hemiptera, Coleoptera, Lepidoptera, Hymenoptera, Diptera, в мае — 10 отрядов. Наиболее часто в ловушки попадали жесткокрылые (42%), двукрылые (18%), равноногие (10%) и пауки (в весеннее время).
В первой половине сентября основное ядро напочвенных беспозвоночных составляют представители отряда двукрылых (семейство Myscidae — настоящие мухи), жесткокрылых (главным образом семейства Tenebrionidae — чернотелки и Carabidae — жужелицы), чешуекрылых (семейства Pterophoridae — пальцекрылки, Puralididae — огневки, Noctuidae — совки), прямокрылых (семейства Acrididae — саранчовые настоящие и Gryllidae — сверчки настоящие).
В первой половине мая герпетобионты представлены в основном отрядами жесткокрылых (главным образом семейства Tenebrionidae — чернотелки и Scarabeidae — пластинчатоусые), перепончатокрылых (семейств Formicidae — муравьи), двукрылых (семейство Myscidae — настоящие мухи) (рис. 1).
Буферная зона является оптимальной средой обитания именно для отряда жесткокрылых. Доля Coleoptera здесь в 2 раза выше по сравнению с другими территориями. Во время весеннего выезда количество учтенных жесткокрылых возросло еще больше, так как на последние месяцы весны и начало лета приходится пик их видовой активности в умеренных широтах. Наибольшее число видов появляется в мае — июне. Также имеется несколько видов, лёт которых приходится на конец летаначало осени. На протяжении вегетационного периода растений происходит постепенная смена фау-нистического состава жесткокрылых, при этом некоторые виды живут в течение практически всего тёплого сезона — с весны до осени.
Из саранчовых характерны: Calliptamus italicus L. — итальянский прус. И личинки, и имаго его сильно вредят пастбищам и сенокосным угодьям. Dociostaurus brenicollis — крестовичка, пустынно-степной вид Dociostaurus kraussi Ing. — атбасарская крестовичка, пустынно-степной обычный вид. Предпочитает полупустынные участки с засоленными почвами и полынной или злаково-эфемероидной растительностью, в основном злаки.
Из других ортоптероидных насекомых на местообитаниях этого биоценоза отмечены представители рода Gryllomorphasp. (Gryllidae).
За счет ортоптероидных насекомых живет большаяупга хищных (скутигеры, тарантулы, пауки-скакуны, богомолы из рода Mantis, Bolivaria, Rivetina, мухи-ктыри и др.) и паразитических насекомых. Из энтомофагов часто встречались перепончатокрылые, особенно Prionyx subfuscatus Dahl. (Sphecidae), обычный на сухих лугах в пустынной зоне. Охотится на саранчовых. В наших сборах преобладали эндопаразитические наездники подсемейств Ichneumoninae, Cryptinae, осы сем. Scoliidae (эктопаразиты личинок пластинчатоусых, долгоносиков) и мухи из сем. Bombylidae, паразитирующие в кубышках саранчовых. На теле саранчовых паразитируют клещи краснотелки (сем. Trombidiidae). Осы Bembixrostrata L. (Crabronidae) своих личинок кормят убитыми крупными мухами (Tabanidae, Syrphidae), а дорожные осы Pompilus viaticus L. (Роmpilidae) — охотники на крупных пауков. Роющие осы охотятся на мелких беспозвоночных.
горнодобывающий беспозвоночный полупустынный антропогенный Под камнями, среди остатков растительности встречаются мокрицы из рода Hemilepistus, скорпионы, многоножки, паразитические клещи, пауки, тарантулы из рода Lycosa. Среди кустов боялыча и курчавки пауки сем. Araneidae (Argiopelobata Aud и др.) строят колесовидные сети и ловят крупных летающих насекомых.
Двукрылые также здесь довольно многочисленны, но предпочитают более открытые и каменистые местообитания, а многие являются потребителями растительных остатков или хищничают. Личинки паразитируют на других насекомых.
Отличительной чертой сезонной встречаемости (сентябрь) было наличие только имагинальных стадий развития для всех групп герпетобионтных беспозвоночных.
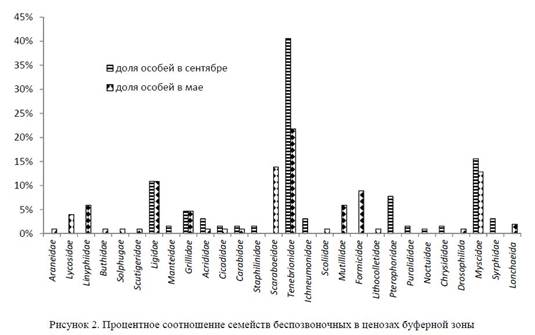
Выявленные отряды были представлены в сентябре 15 семействами: Linyphiidae, Ligidae, Manteidae, Grillidae, Acrididae, Cicadidae, Carabidae, Staphilinidae, Tenebrionidae, Pterophoridae, Puralididae, Ichneumonidae, Chrysididae, Myscidae, Syrphidae. Преобладают представители семейства чернотелок Tenebrionidae (41%), сопутствующими являются Myscidae (16%), Ligidae (11%). В мае количество семейств увеличилось до 28. Наиболее многочисленным семействами с мая по сентябрь постоянно оставались чернотелки. Семь семейств, представляющих фито и сапрофагов, регулярно регистрировались здесь и весной, и осенью: Ligidae, Grillidae, Acrididae, Cicadidae, Carabidae, Tenebrionidae, Myscidae (рис. 2).
Среди жесткокрылых в буферной зоне явно доминируют жуки-чернотелки. По характеру питания они относятся к ярко выраженным фитосапрофагам. Их имаго на поверхности почвы поедают степную подстилку, личинки — мертвые корни.
В сухих районах некоторые виды сильно вредят культурным растениям и пастбищам, повреждают пищевые запасы, немногие хищны. Кроме того, это организмы — минерализаторы, которые производят глубокую переработку опада, препятствуя его трансформации в гумус, сдвигая его в сторону минерализации. Кроме непосредственного поедания подстилки, чернотелки разлагают подстилку, измельчая ее и увеличивая тем самым поверхность разлагающегося субстрата. Экскременты чернотелок — центры стимуляции целлюлозоразрушающей микрофлоры, что дополнительно усиливает разложение подстилки. Среди ценотических типов умеренного климатического пояса Азии полупустыни, пустыни и степи имеют самый низкий общий запас растительного вещества, очень большую продукцию фитомассы и беспрецедентно высокую скорость её деструкции. Поэтому чернотелки, с их количественным потенциалом и функциональными возможностями, вносят значительный вклад в эти процессы.
В ряду зональных степных и пустынных биогеоценозов уровни численности и биомассы чернотелок увеличиваются от северного предела степей к сухим, пустынным и полупустынным, после чего снова уменьшаются. От влажных вариантов степей и полупустынь к сухим растет число особей, доминантные позиции переходят от Crypticini, Opatrini, Pedinini к Blaptini, Platyscelidini и, наконец, к Tentyriini.
Для естественных ландшафтов буферной зоны характерна максимальная уловистость чернотелок. Она примерно в 2 раза выше, чем для других территорий, составляя U = 0,7/0,8 (жуки чернотелки/жуки). Здесь же встречено и максимальное количество видов, принадлежащих к пяти родам: Tentyrianomas (Pallas, 1781); Pimeliainterpunctata Klug, 1830; Adesmiaanomala (F.-W., 1820); Pimeliacephalotes (Pallas, 1781); Blapsdeplanata Menetrie, 1832; Blapspruinosa Faldermann, 1833.; Opatroidespunctulatus Brulle, 1832.
Самыми массовыми оказались представители родов Pimelia и Blaps. Возможно, это свидетельствует о том, что биотопические условия на этой территории сохранили в наибольшей степени черты природной зоны и изменены не столь значительно. Чернотелки из рода толстяков (Pimelia и близкие роды) — крупные чернотелки с массивным, округлым, более или менее выпуклым телом и часто с хорошо развитыми на ногах щетками из длинных волосков, которые облегчают передвижение по песку и закапывание в него. Вместе с ними в биоценозах многочисленны более мелкие, быстро бегающие, похожие на жужелиц тентирии (Tentyria и близкие роды). Виды рода Adesmia — обитатели пустынь, отличающиеся необычайно длинными ногами.
Tenebrionidae имеют разнообразные морфо-физиологические адаптации к условиям жизни в аридных регионах. Так, смещение времени активности на утренние, вечерние и ночные часы является одним из важных моментов их биологии, позволяя существовать и процветать в крайних условиях пустынь. Среди крупных форм встреченных нами чернотелок (сумеречные и ночные формы) может быть выделена группа типа Blaps. Они редко встречаются при проведении дневных маршрутных учетов. Среди дневных форм выделяется группа типа Pimelia.
В целом же, с увеличением засушливости, чернотелки становятся главным агентом форсированной минерализации растительных остатков в пустынях и степях с коротким вегетационным периодом. Уменьшение числа или исчезновение видов чернотелок, являющихся эдификаторами состояния аридных территорий, приведет к непоправимому нарушению состояния пустынных биоценозов.
Характерной чертой весеннего периода стало появление в сборах достаточно многочисленной (15%) группы пластинчатоусых жуков (Scarabeidae), представленной главным образом копрофагами из рода Onthophagus (рис. 2). Эвритопный ксерофил, предпочитающий сухие участки, Opatrum sabulosum L. относится к многоядным вредителям. Жуки повреждают весной всходы злаков, у проросших растений объедают семядоли и молодые листочки около поверхности почвы, подгрызают и перегрызают стебельки молодых растений у основания. Личинки повреждают семена, корни, подземные части стеблей различных культур.
Общая динамика плотности герпетобионтных беспозвоночных в буферной зоне составила 1,8 экземпляра на ловушко/сутки. При этом максимум уловистости приходится на долю жесткокрылых (Coleoptera — 0,8 экз. на ловушко/сутки), почти в три раза меньше уловистость двукрылых (Diptera — 0,3 экз. на ловушко/сутки). Несколько ниже уловистость мокриц и чешуекрылых (Isopoda и Lepidoptera — 0,2 экз. на ловушко/сутки), а также прямокрылых и перепончатокрылых (Orthoptera — 0,13 и Hymenoptera — 0,1 экз. на ловушко/сутки). На долю паукообразных, богомолов и клопов (Aranei, Mantoptera, Hemiptera) приходится самая маленькая уловистость в этом биоценозе, составляющая 0,03 экз. на ловушко/сутки (рис. 3).
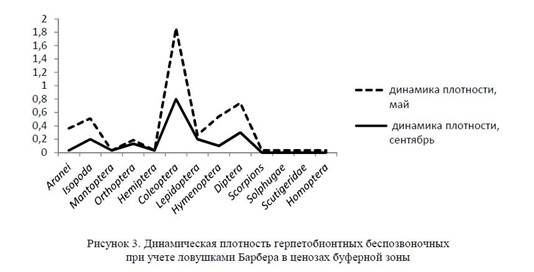
Чаще, чем на других участках, на буферной территории встречались пустынные мокрицы U = 0,2, свидетельствуя о большем разнообразии микроусловий, в том числе создаваемых растительностью.
В мае общая динамическая плотность составила 2,8 экземпляра на ловушко/сутки. Максимум уловистости приходится на долю жесткокрылых (Coleoptera — 1,6 экз. на ловушко/сутки), почти в четыре раза меньше уловистость двукрылых и препончатокрылых (Diptera, Hymenoptera — 0,44 экз. на ловушко/сутки). С сентября по май для пустынных сапрои фитофагов — равноногих рачков, жесткокрылых и двукрылых — регистрировались максимумы динамической плотности. В мае пики плотности превышали осенние показатели примерно в три раза.
Общая трофическая специализация всех поверхностнообитающих беспозвоночных в биоценозах буферной зоны была представлена следующим образом: 73,4%, — фитофаги; 15,6% — сапрофаги; 6,3% — хищники и 4,7% энтомофаги (рис. 4). Из фитофагов преобладает семейство Tenebrionidae, из хищников — жуки-жужелицы и паукообразные. Обитающие здесь фитофилы в своем развитии и питании связаны с растениями-доминантами: боялышем, полынью, тамариском и др. Популяции их немногочисленны, но в отдельные годы развиваются обильно (например, боялышевая совка Odontelia arbusculae Sukh.).
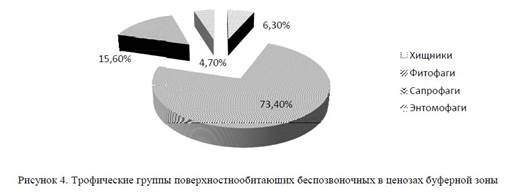
Сапрофагами являлись, в основном, двукрылые семейства Myscidae. Большая часть входящих в это семейство видов развивается в гниющих органических остатках растительного и животного происхождения, где их личинки либо перерабатывают сами остатки, либо хищничают.
За счет ортоптероидных насекомых живет большая групга хищных (скутигеры, тарантулы, пауки-скакуны, богомолы из рода Mantis, Bolivaria, Rivetina, мухи-ктыри и др.) и паразитических насекомых. Значительная часть энтомофагов — наездники. На стадии личинки они — паразиты личинок и куколок многих групп насекомых, в том числе сельскохозяйственных вредителей. Обычными хозяевами ихневмонид являются чешуекрылые, перепончатокрылые (пилильщики, осы, наездники), двукрылые, жуки, реже сетчатокрылые, верблюдки, ручейники. Некоторые виды паразитируют на пауках и ложноскорпионах, соответственно преобладают в аридных зонах. Наездники нуждаются в ежедневном потреблении воды, в связи с чем они могут попадать в ловушки, наполненные жидкостью. Многие виды ведут сумеречный или ночной образ жизни. В наших сборах преобладали эндо-паразитические наездники подсемейств Ichneumoninae, Cryptinae, Scoliidae.
Из хищников встречены паукообразные. Пауки из семейства Linyphiidae некрупные, строят сеть в виде горизонтального навеса или балдахина из плотной паутинной ткани, который растянут на многочисленных вертикальных и перекрещенных нитях [5].
В целом выявленная на данном этапе фауна насекомых и других беспозвоночных исследуемой зоны достаточно разнообразна и характерна для полупустынных растительных сообществ. Доминантной группой герпетобионтов естественных ландшафтов района исследования является семейство Tenebrionidae (41−44% общей численности). Фоновые группы: Myscidae (13−16%), Ligidae (11%), Grillidae, Acrididae (7,8%). Обнаруженные виды чернотелок относятся к Северо-Туранской, Ирано-Туранской и Казахстанской группе. Жесткокрылые и прямокрылые — типичные обитатели пустынь и полупустынь, ксерофилы и фитофилы (доля в трофической структуре 73%, 8−12 семейств); в своем развитии и питании связаны с растениями-доминантами: боялышем, полынью, курчавкой и др. Трофическая структура комплекса характерна для аридной зоны. Доля хищных и паразитических насекомых в трофической структуре — 6,3%. Полученные данные по фоновой территории могут служить основой для прогнозирования изменений пустынных биоценозов, поддержания оптимума экобаланса в них и проведения мероприятий, необходимых для защиты и сохранения видов пустынных регионов Северного Прибалхашья.
- 1. Ашихмина Т. Я. Биоиндикация и биотестирование — методы познания экологического состояния окружающей среды. — Киев: Наук. думка, 2005. — 246 с.
- 2. Чигаркин А. В. Региональная геоэкология Казахстана. — Алматы: ?аза? ун-ті баспасы, 2000. — 224 с.
- 3. ГвоздецкийA., Николаев B.A.Казахстан. — М.: Мысль, 1971. — 296 с.
- 4. ГиляровМ.С. Методы почвенно-зоологических исследований. — М.: Наука, 1975. — 280 с.
- 5. Крыжановский О. Л. Состав и распространение энтомофаун земного шара. — М.: КМК, 2002. — 237 с.