Прибрежно-водная растительность водотоков (горных ручьев и рек)

Верхняя часть склонов долины горных ручьев обычно имеют V-образный поперечный профиль, а само русло водотока — форму лотка. Ширина русла обычно не превышает одного метра, а глубина редко бывает более полуметра. На этом участке водотока на протяжении до нескольких сот метров (в зависимости от литологии горной системы) грунт русловой части представляет собой нагромождение крупноглыбового материала… Читать ещё >
Прибрежно-водная растительность водотоков (горных ручьев и рек) (реферат, курсовая, диплом, контрольная)
Характер развития прибрежно-водной растительности в озерах, водохранилищах, прудах описан в предыдущих главах. В этих водоемах распределение растительности носит в основном зональный характер, присущий тому или иному типу водоема и зависящий от многих физико-химических, гидрологических и биотических факторов.
В настоящей главе представлены сведения о растительности водотоков (горных ручьев, равнинных рек и прирусловых водоемов), в которых развитие растений в большей степени зависит от мощности паводка, скорости течения и характера аллювиальных отложений (П. С. Кузин, 1960).
Несмотря на ряд сходных черт и общих закономерностей в формировании сообществ, прибрежно-водная растительность в бассейне каждого водотока имеет индивидуальные особенности, отличаясь флористическим составом, занимаемой площадью и пространственным распределением. Описать все эти особенности не представляется возможным, поэтому мы это делаем на примере некоторых наиболее характерных водотоков Дальнего Востока с присущей им растительностью (М. А. Кудряшов, 1977).
В горных биотопах встречаются различные группы наземных растений — мезофиты в самом широком понимании. Они объединены в группу «прибрежной растительности» приуроченностью произрастания в прирусловой части водотока на постоянно трансформирующемся грунте. В распределении этих растений наблюдается определенная закономерность. В прирусловой части горных ручьев при постоянной эрозии грунта сообщества растений состоят из гигро-, мезофитных психрофильных видов. Вниз по течению водотока в зоне аккумуляции аллювия разного размера, в условиях водного дефицита (в период между паводками) и достаточной освещенности, встречаются и ксеромезофитные растения. Необходимо отметить, что подобного рода водотоки характеризуются резко меняющимся гидрологическим режимом — от бурных потоков во время паводков до сухого русла в период межени.
К прибрежно-водной растительности горных биотопов можно отнести растения, обитающие в таких зонах динамичного гидрологического режима водотока, как зоны эрозии, транзита и переотложения аллювия. Эти сообщества по флористическому составу сильно отличаются от растительности окружающих территорий. Галечники с их пионерной растительностью раскрывают картину развития пойменной растительности. Поселившись на галечниках, эти растения дают начало растительному покрову поим в зоне активного аллювиального процесса (М. А. Кудряшов, 1988).
Верхняя часть склонов долины горных ручьев обычно имеют V-образный поперечный профиль, а само русло водотока — форму лотка. Ширина русла обычно не превышает одного метра, а глубина редко бывает более полуметра. На этом участке водотока на протяжении до нескольких сот метров (в зависимости от литологии горной системы) грунт русловой части представляет собой нагромождение крупноглыбового материала, а также выходов коренных пород, создающих специфические уступы. Русло на этом отрезке обычно сухое, и только в период паводков под валунами течет вода. Поверхностный сток в период межени может идти на глубине нескольких метров. Отсутствие субстрата, пригодного для роста растений на этом участке русла, не позволяет говорить о наличии здесь какой-либо растительности.
По мере увеличения площади водосборного бассейна происходит дальнейшее развитие долинного рельефа; появляется выраженное русло, которое все глубже и шире врезается в днище долины. В дальнейшем размеры долин возрастают и достигают ширины 30—50 м и более. Поперечный профиль долин становится корытообразным, с достаточно крутыми бортами, а грунт характеризуется повышенным содержанием мелкозема (А. И. Кичигин, 1974). С появлением тонкообломочного материала в русловой части горного ручья отмечается появление воды. Это сопровождается формированием растительных сообществ, характерных для аллювия из песчано-суглинистого и галечного материала, условий повышенной влажности и своеобразного температурного и светового режима долины.
Для иллюстрации описанного выше в качестве примера изображены четыре схемы поперечных профилей горного ручья в бассейне реки Зеркальная (Сихотэ-Апинь) через каждые 200 м (рис. 13).
Первый профиль, заложенный при появлении выраженного русла, пока лишен растительности. Русло сложено крупноглыбовым материалом, непригодным для произрастания растений. Второй профиль заложен на месте поступления мелкообломочного материала. Здесь уже встречаются характерные растения группы формаций прирусловых мезофитов. Вниз по течению по мере увеличения доли песчано-глинистой фракции видовой состав прибрежных мезофитов расширяется.
Размещение растений в прирусловых сообществах имеет мозаичный характер, обусловленный неоднородностью механического состава субстрата и особенностями его распределения. Сообщества, произрастание которых приурочено к прирусловой части горных ручьев, относят к «прибрежной растительности», они входят в нее в виде особой группы формаций прирусловых мезофитов.
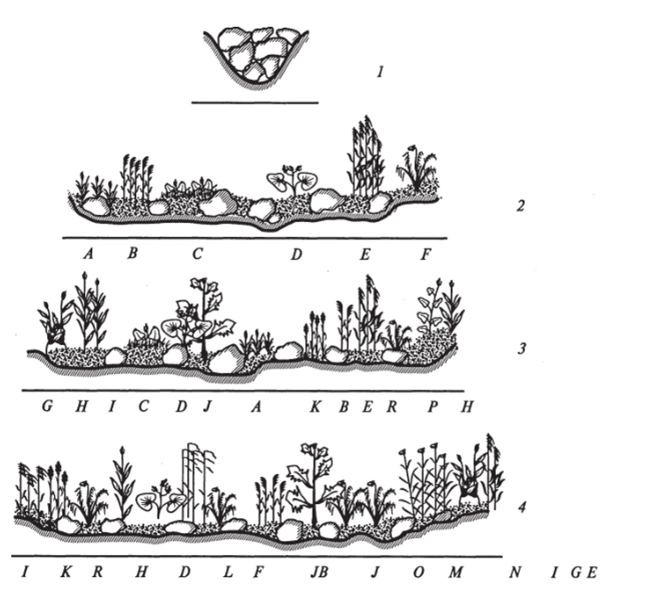
Рис. 13. Поперечные профили (1—4 1 — растения отсутствуют) растительности горного ручья в бассейне р. Зеркальная:
А — Carex uda; В — Digraphis arundinacea; С — Chrysosplenium Jlagel! iferum D — Callha silvestris; E — Urtica anguslifolia; F — Anthyrium rubripes', G — Cardamine yezoensis; H — Artemisia rubripes; /— Impatiens noli-tangere; J — Filipendula palmate', К — Epilobium cylyndrostigma', L — Aconitum axilliflorum', M — Anthriscus aemula; N — Gliceria lithuanica; О — Dryopteris buschiana', P — Cacalia hastate', R — Athyrium
coreanum.
Масштаб 1 см: 0,6 м В этих растительных сообществах выделяют четыре группы по биологическим особенностям, позволяющим им переносить паводковый режим (М.А. Кудряшов, Е. Б. Поспелова, 1981; М. А. Кудряшов, А. П. Садчиков, 2002).
Первую группу представляют многолетники с хорошо развитой корневой системой, проникающей в толщу аллювиальных отложений на глубину более 50 см: калужница (Callha), воронец (Actaea), шпороцветник (Aconitum) и др. Эта группа растений переносит паводок за счет заглубленной и хорошо разветвленной в толще аллювия корневой системы, которая не повреждается током воды. Надземная часть растений, разрушающаяся практически полностью во время паводка, восстанавливается в течение двухтрех недель.
Ко второй группе относят растения, тяготеющие в произрастании к мелкогалечному аллювию с повышенным содержанием песчаной фракции: цинна (Cinna), сердечник (Cardamine), двукисточник (Phalaroides = Digraphis), овсяница (Festuca) и многие другие. Они способны к вегетативному возобновлению из погребенных паводком в толщу грунта частей растений. Паводок является для них и фактором расселения: поток воды переносит части растений, которые на новом месте продолжают вегетировать.
Произрастание третьей группы растений приурочено к наносам самых мелких фракций: виды селезеночника (Chrysosplenium), фрима (Frima), двулепестник (Circea) и др. Эти виды удерживаются в прирусловой части за счет куртинного характера роста, способности к интенсивному вегетативному размножению и плодоношению до наступления периодов паводков. Благодаря поверхностно расположенной корневой системе они легко смываются паводком и переносятся куртинами по прирусловой части ручья.
К четвертой группе относятся однолетние растения, которые произрастают в прирусловой части ручья благодаря высокой семенной продуктивности, в частности виды недотроги (Impatiens). Постоянная эрозия грунта в прирусловой части ручьев обуславливает пространственное изменение в размещении отдельных видов этой группы растений до и после паводка.
Каждая из перечисленных групп растений обладает различной подвижностью в пределах прирусловой части ручьев. Наименее подвижны виды первой группы растений с развитой корневой системой. Они переносятся только во время паводка вместе с передвигающимся слоем аллювия. Другие группы растений перемещаются по руслу ручья во время паводка более интенсивно.
На участках прирусловой части ручья, сложенных аллювием с преобладанием мелкообломочной фракции, наиболее характерно трехъярусное распределение растений. В первом ярусе обычно располагаются такие виды, как лабазник (Filipendula), крапива (Urtica), шпороцветник (Aconitum), недотрога (Impatiens). Второй ярус составляют кочедыжник (Athyrium), калужница (Caltha), сердечник (Cardamine), цинна (Cinbna), двукисточник (Digraphis). Третий ярус формируют селезеночник (Chrysosplenium), двулепестник (Circea) и др. Следует отметить, что такое строение сообщества с выраженными ярусами наблюдается только в середине вегетационного периода. Для весенней фазы развития характерно двухъярусное строение.
В зависимости от климатических особенностей в сообществе наблюдается различное количественное соотношение видов. В засушливые периоды растения с поверхностной корневой системой развиваются слабее по сравнению с видами, имеющими более мощную корневую систему.
Флора прирусловой части горных ручьев достаточно однородна для многих водотоков. При рассмотрении видовой насыщенности прирусловых растительных сообществ наблюдается прямая зависимость от присутствия в наносах мелкодисперсных фракций аллювия. По мере развития долинного рельефа прирусловая часть расширяется, а мелкий и влажный грунт улучшает условия произрастания растений. За счет этого видовой состав сообщества приближается к максимально возможному, а, как известно, высокое постоянство флористического состава свойственно хорошо сформированным и устойчивым сообществам (А. П. Шенников, 1964).
По мере выхода горных ручьев и рек в долины активность водной эрозии ослабевает. Русловой аллювий, вынесенный из верховьев, начинает откладываться вдоль потока, образуя береговую отмель, которая после каждого паводка увеличивает свои размеры в длину и высоту. На распределение гальки и более мелких фракций наноса влияет рельеф водотока. Ближе к берегу откладывается более тяжелая фракция — крупная галька, а далее от него — более легкие фракции. Иногда во время сильного паводка часть участка размывается и уносится потоком. Таким образом, в пойме происходит постоянный процесс накопления и разрушения аллювия. В геоморфологическом отношении такая прирусловая отмель представляет собой зачаточную часть поймы (А. П. Нечаев, 1960); прирост пойменного аллювия составляет около 1 см/год (Н. Г. Васильев, 1962).
Своеобразие растительности речных пойм проявляется в том, что ее флора содержит элементы, не свойственные растительности окружающего водораздела. В то же время водный режим формирует сходные сообщества пойм, находящихся в разных географических зонах. Несмотря на существование ряда сходных сообществ, характерных для пойм рек и наличия общих закономерностей в размещении ценозов по поперечному профилю долины, растительность каждой поймы своеобразна. Она различается количеством представленных сообществ, соотношением их площадей, флористическим составом. Большое число факторов, определяющих тип условий экотопа (эдафических, гидрологических, литологических и др.), обуславливает развитие тех или иных сообществ (А. П. Шенников, 1950; Р. А. Еленевский, 1938).
Особое значение эти факторы имеют для развития сообществ на отмелях рек Дальнего Востока, которые мы взяли в качестве примера. Их растительность, несмотря на ряд общих черт с отмельной растительностью других географических зон, все же отличается своей флористической своеобразностью. Прирусловую отмельную растительность быстрых рек подразделяют на две группы.
Первая группа растительного сообщества приурочена к галечным отмелям, сложенным в основном галькой и гравием. Песчаная и глинистая фракции здесь незначительны. Уровень воды находится в сфере досягаемости корневых систем растений. Широкая амплитуда абиотических факторов позволяет произрастать здесь значительному числу видов растений. Это различные виды злаков — щетинник зеленый (Setaria viridis), лисохвост амурский (Alopecurus amurensis), полевицы (Agrostis alba, A. clavata), мятлик луговой (Роа pratensis), горцы (Polygonum foliosum, Р. sieboldii), осоковые (Сагех vesicata, Cyperus amuricus, С. truncatus).
Вторая группа растительности приурочена к песчаным наносам в районе равнинных расширений, которые наряду с песком содержат глинистые и илистые частицы. Кроме однолетних растений, таких как щетинник (Setaria), гулявник (Sisimbrium), росичка (Digitaria), в этой зоне произрастают и многие многолетники — некоторые горцы (Polygonum), смолевки (Silene), лапчатки (Potentilia).
Во внешней части песчаных и песчано-илистых отмелей только небольшое число видов могут образовывать сомкнутые ценозы, которые можно рассматривать как группы ассоциаций: череды (Alisma orientate), болотницы (Eleocharis kamtschatica), триостренника (Triglochin maritimum), торичника (Spergularia salina), гляукса-млечника (Glaux maritima).
Во внутренней зоне песчаных отмелей многолетние осоковые и злаковые растения образуют моноценозы, располагающиеся в виде поясов или куртин. Поэтому представляется возможным выделение групп ассоциаций следующих растений: вейника (Calamagrostis angustifolia Кош., Cal. langsdorffii Trin.), осоки (C. cryptocarpa С. A. Mey., C. kirganica Кот., C. lasiocarpa Ehrh., C. mollissima Christ., C. pseudo-curaica F.Schmidt., C. neurocarpa Maxim., C. rhynchophysa C.A.Mey., C. schmidtii Meinch., C. tuminensis Kom., C. vesicata Meinch.), хвоща (Equisetum limosum L.), болотницы (Eleocharis mamillata Lindb.), манника (Glyceria spiculosa (Fr. Schmidt.) Roshev.), тростника (Phragmites communis Trin.), горца (Polygonum amurense Niew.), лапчатки (Potentilla pacifica Howell.), стрелолиста (Sagittaria natans Pall.), камыша (Scirpus compactus Hofim., Sc. tabemaemontani C.C.Gmel.), ежеголовника (Sparganium glomeratum Laest., Sp. stenophyllum Maxim.), рогоза (Typha latifolia L.).
Расположение поясов и переходных зон между ними изменяется во времени и пространстве по площади отмели, что объясняется постоянным изменением условий обитания, в основном под действием паводка. В качестве примера приводим изменение расположения поясов растительности на песчаных отмелях реки Зеркальная в течение трех лет (рис. 14).
При первом наблюдении во внешней зоне отмели шириной около 5 м отмечена агломерация из 11 видов однолетников. Ближе к внутренней зоне располагается пояс частухи восточной (Alisma orientate (Sam.) Juz.) шириной около 2,5 м. Внутренняя зона начинается с пояса рогоза широколистного (Typha latifolia L.) шириной 2 м, второй пояс — это пояс тростника обыкновенного (Phragmites communis Trin.) шириной 3 м, третий пояс — вейника узколистного (Calamagrostis angustifolia Кош.) шириной более 4 м.
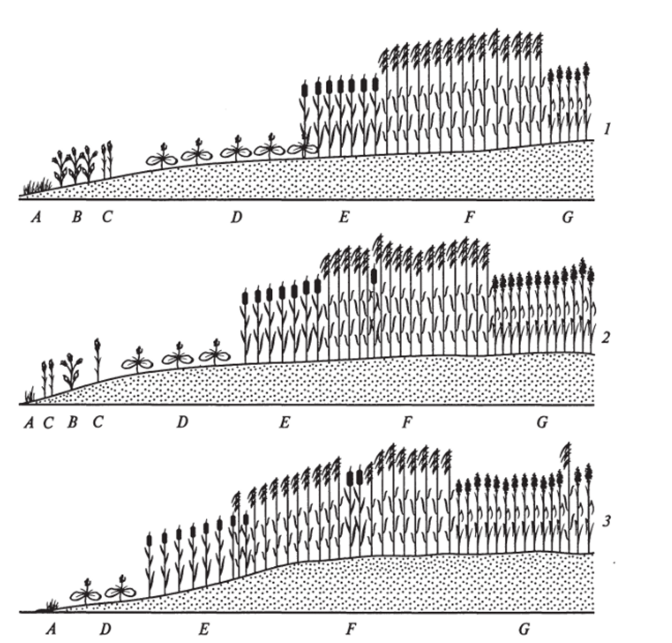
Рис. 14. Изменение расположения поясов растительности на песчаной отмели р. Зеркальная:
А — Gnaphalium uliginosum; В — Bidens cernua; С — Setaria viridis; D — Alisma orientate; E— Typha latifolia; F— Phragmites communis', G— Calamagrostis angustifolia.
Масштаб 1 см: 1 м Между этими поясами наблюдались переходные зоны различной ширины с выраженным взаимопроникновением растений.
Через год наблюдался сдвиг зон растительности в сторону внешней части отмели. Пояс рогоза сдвинулся на внешнюю часть отмели. Через два сезона после сильного паводка почти вся внешняя зона песчаной отмели была смыта. На узкой полосе, оставшейся от внешней зоны, остались единичные экземпляры частухи, горца, череды, сушеницы. Паводок, смывший большую часть внешней зоны, не нанес существенного ущерба поясам многолетников — рогозу, тростнику, вейнику. Такие процессы сдвигов поясов произрастания растений проходят в пойме постоянно.
Наиболее быстро возникают и разрушаются сообщества во внешней зоне отмели, возле уреза воды. Сообщества внутренней зоны отмели более устойчивы из-за развитой системы подземных побегов, армирующих толщу отмели. Тростник быстрее всего захватывает новые аллювиальные наносы, образовывая за вегетационный период надземные стелющиеся побеги длиной более 8 м. Побеги укореняются в узлах и дают начало новому клону. Отдельные части таких побегов могут переноситься потоком и дают начало новой колонии тростника. Тростник, задерживая своими зарослями аллювиальные частицы, повышает уровень отмели, что приводит к смене моноценозов тростника моноценозами вейников и осок. На защищенных от воздействия паводка участках отмели развиваются ценозы манника колоскового, горца амурского, стрелолиста плавающего, ежеголовников. Корневая система этих растений недостаточно заглублена в грунт и паводок наносит им значительный ущерб. Однако они хорошо приспособлены к колебанию уровня воды и в защищенных от потока участках реки могут заходить в воду, образуя водные формы. Особенно отчетливо проявляется способность к освоению водной среды у таких прибрежных растений, как горец амурский и стрелолист плавающий.
Флора и растительность отмелей рек Дальнего Востока изучалась многими исследователями (А. П. Нечев, 1960; В. Н. Ворошилов, 1968; 3. И. Гапека, 1971, 1974). Кроме того, существует ряд работ, посвященных флоре и растительности отмелей рек в других регионах нашей страны: Камы (К. Н. Игошина, 1927), Волги (М. В. Марков, 1950), Урала (В. М. Пешкова, 1950), Енисея (Л.И.Номоконов, 1959), Мета (И. В. Самойлов, 1970) и др.
Многие реки и прирусловые водоемы Дальнего Востока подвержены регулярным колебаниям уровня воды. Из-за этого некоторые виды гидатофитов приспособились к временному пересыханию водоемов, образуя наземные формы, которые могут некоторое время находиться вне водной среды. Среди таких видов можно назвать водяную сосенку (Hippuris vulgaris L.), уруть колосистую (Myriophyllum spicatum L.), болотноцветник щитовидный (Nymphoides peltata (S. G. Gmel.) O. Kuntze). Однако большинство гидатофитов, произрастающих на глубинах в пределах 0,5 — 2 м, не переносят пересыхание водоема. При резком понижении уровня воды до 0,1—0,2 м отмирают многие массовые виды, и в первую очередь водяной орех (Тгара incisa Sieb. et Zucc., T. mandshurica Fler.), рдест курчавый (Potamogeton crispus L.), кувшинка (Nymphaea tetragona Georgi). Регулярные колебания уровня воды в реках приводят к образованию между прибрежной растительностью и водой полосы, лишенной растительности. Ее ширина зависит от амплитуды колебания уровня воды в реках.
Сообщества водных растений по строению и составу сложены более просто, чем сообщества прибрежных растений. Это связано с тем, что большинство гидатофитов произрастает в виде монодоминантных колоний. В частности, это рдесты — Potamogeton pusillus L. и Р. perfoliatus L. (3. И. Гапека, 1971, 1974); виды кувшинок (Nymphaea), кубышек (Nuphar), наяды (Najas), водяного ореха (Тгара), роголистника (Ceratophyllum) и др. Заселение текучих вод растительностью происходит в основном за счет укоренения и размножения вегетативных побегов, однако не исключено заселение русла реки и семенным путем (Г. Вальтер, 1975).
Колонии гидатофитов обильно разрастаются в слабопоточных и мелководных протоках рек. Илистое дно проток, более высокая температура воды в них по сравнению с основным руслом, меньшие скорости течения во время паводков — все это позволяет гидатофитам образовывать здесь заросли с проективным покрытием до 90%.
Стоячие водоемы поймы рек представлены многочисленными старицами, озерами и протоками, потерявшими связь с основным руслом реки, однако во время паводка или разлива они соединяются с рекой. Площадь водного зеркала и глубина таких водоемов различна. Во время летне-осенних паводков вся пойма покрывается водой на десять—двенадцать дней. Создаются благоприятные условия для переноса проростков водных растений, поэтому их видовой состав близок для водоемов данной территории (М.П. Шилов, 1972).
В стоячих и слабопроточных водоемах можно наблюдать отчетливое поясное распределение водной растительности (рис. 15). Первая зона, следующая сразу за полосой прибрежных растений, представлена формациями наводноплавающих укореняющихся растений, произрастающих на глубинах до 0,7 м: кувшинки (Nymphaea), кубышки (Nuphar), водяного ореха (Тгара), болотноцветника (Nymphoides). Вторая зона образована формациями подводноплавающих укореняющихся растений, распространенных на глубинах от 0,7 до 1 м. Здесь произрастают рдесты (Potamogeton), уруть (Ceratophyllum), перистолистник (Myriophyllum), наяда (Nayas), водяная сосенка (Hyppuris). Третья зона представлена формациями.
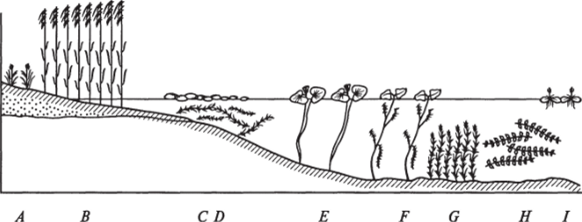
Рис. 15. Поясное распределение прибрежно-водной растительности в слабопроточном водоеме:
А — Carex leiorhyncha', В — Scirpus tabernaemontani С — Ceralophyllum oryzetorum; D — Lemna minor; E — Nymphaea lelragona', F — Trapa incise; G — Potamogeton peifolialus; H — Myriophyllum spicalum; I — Ulricularia intermedia
неукореняющихся плавающих гидатофитов: ряски (Lemna) и многокоренника (Spirodela), которые перемещаются в зависимости от ветровых условий по всей поверхности водоема. Эти растения могут входить в колонии любой зоны или образовывать изолированные группы.
При эвтрофировании стоячих водоемов создаются условия для массового развития плавающих неприкрепленных растений — сальвинии (Salvinia), пузырчатки (Ulricularia), вольфии (Wolffia), альдрованды (Aldrovanda). Как «естественные светофильтры» они препятствуют проникновению света в толщу воды и тем самым вызывают отмирание погруженных растений. Это приводит к дальнейшему обогащению воды органическим веществом (Г. Вальтер, 1975). Развитие процесса приводит к наступлению прибрежной растительности на обмелевшие участки, что вызывает отмирание плавающих растений из-за недостатка света.
Устьевая часть рек подвержена засолению морскими водами. Это сказывается на флористическом составе и распределении растений. Проникновение морских вод в устья рек приводит к изменению отмельных сообществ. Меньше всего подвержены действию солей сообщества тростника (Phragmites communis Trin.) и камыша (Scirpus tabemaemontani С. C.Gmel.), наиболее устойчивые к повышенному содержанию солей в воде и грунте (до 25%о). Они образуют на засоленных отмелях сообщества, представленные монодоминантными зарослями.
По мере приближения к устью и увеличения засоленности происходит угнетение зарослей тростника и камыша. На илисто-глинистых или илисто-песчаных отмелях появляются типичные галофитные виды: глаукс-млечник приморский (Glaux maritima L.), триостренник морской (Triglochin maritimum L.), торичник морской (Spergularia salina J. et C.Presl.), болотница камчатская (Eleocharis kamlschatica (С. A. Mey.) Кош.), солянка Комарова (Salsola komarovii Iljin). В прилив эти сообщества или группировки часто покрываются слоем воды до 30 сантиметров.
На песчано-галечном субстрате сообщества или группировки представлены видами, характерными для морских побережий: осока Гмелина (Carex gmelinii Hook, et Аш.), полынь Стеллера (Artemisia stelleriana Bess.), льнянка японская (Linaria japonica Miq.), мертенсия приморская (Mertensia maritima (L.) S.F.Gray.). Сообщества имеют вид полос или пятен вдоль как основного русла, так и проток и озер. На участках, подверженных воздействию приливноотливного цикла, примыкающих непосредственно к литорали, встречаются морские галофиты: зостера морская (Zostera marina L.) и филлоспадикс иватензе (Phyllospadix iwatensis Makino).