Нейрофизиологические основы распознавания
Нейрофизиологические данные о потоках в обработке зрительной информации свидетельствуют в пользу того факта, что распознавание зрительного паттерна происходит одновременно в нескольких направлениях — от выделения в нем простейших элементов (определяющих контуры объекта, а следовательно и его форму) до целостного — структурного — сличения. В пользу последнего свидетельствуют данные о модульной… Читать ещё >
Нейрофизиологические основы распознавания (реферат, курсовая, диплом, контрольная)
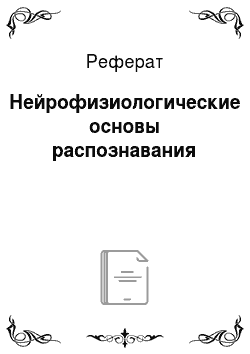
В последнее время психологическая наука все чаще обращается к нейрофизиологическим данным для объяснения различных психических явлений и феноменов. Так, открытие Хьюбелом и Визелом нейронов-детекторов в зрительной коре кошки составило нейробиологический фундамент для теорий признаков. Исследование более высоких уровней в работе зрительной системы также значительно обогатило современную психологическую науку данными о механизмах распознавания паттернов. В 1982 г. два американских ученых Лесли Анджерлейдер и Мортимер Мишкин описали эксперимент, результатом которого стало открытие двух относительно независимых путей, или потоков, в обработке зрительной информации, обслуживающих разные функции восприятия[1]. Под путем в современной нейрофизиологии понимается направленное движение нервных импульсов к определенным нервным центрам. Мишкин и Анджерлейдер предъявляли макакам две задачи на распознавание: задачу на распознавание объекта и его формы и задачу на распознавание пространственных отношений фигур (иначе — ландшафта). В первом случае животному предъявляли два объекта различной формы (например, параллелепипед и призму), за выбор одного из объектов животное поощрялось (за правильной фигурой располагалось угощение); такая схема привела к тому, что при предъявлении положительно подкрепляемой фигуры с какой-либо другой, животное всегда выбирало первую. Во второй задаче макаке предъявлялись три фигуры — две плоские пластины и цилиндр; одна пластина располагалась ближе к цилиндру, другая — дальше. Животное обучали за счет вознаграждения убирать ту фигуру, которая была расположена ближе к цилиндру (рис. 8.21). Мишкин и Анджерлейдер удаляли или разрушали небольшие области мозга макаки, участвующей в обучении, и следили, какие функции восприятия при этом нарушались. После удаления височной доли животные не могли решать задачу по распознаванию объекта: предъявление двух фигур, выбор одной из которых раньше поощрялся, не приводило к положительному эффекту. Мишкин и Анджерлейдер назвали путь, идущий от зрительной зоны VI в нижнюю височную долю системой «Что?», или вентральным потоком. Эта затылочно-височная система ответственна за распознавание объекта, т. е. позволяет ответить на вопрос «Что это за объект?».
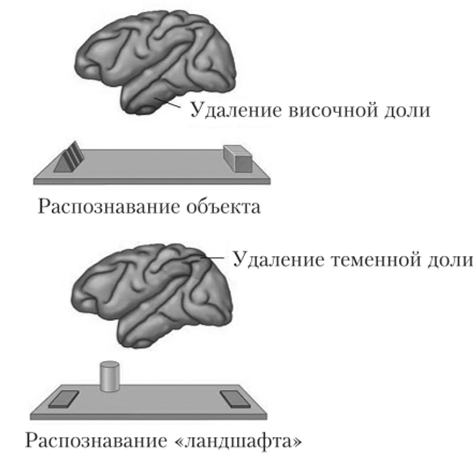
Рис. 8.21. Различные задачи и участки мозга, обеспечивающие успешное их решение, в эксперименте Мишкина и Анджерлейдер1.
После удаления задней части теменной доли животное испытывало затруднение с решением второй задачи — на распознавание пространственных положений объектов. Этот результат позволил выделить путь, идущий из затылочной доли в теменную, который авторы назвали системой «Где?», или дорсальным потоком. Эта система ответственна за определение расположения объекта внешней среды и отвечает на вопрос «Где расположен объект?» Связи и направления потоков показаны па рис. 8.22. Оба пути не являются абсолютно независимыми и однонаправленными. Дальнейшие опыты Анджерлейдер с коллегами показали, что высшие корковые центры посылают сигналы обратно в зону VI, что, с одной стороны, составляет систему обратной связи, а с другой стороны, представляет собой нейрофизиологический базис «нисходящих процессов» переработки[2][3].
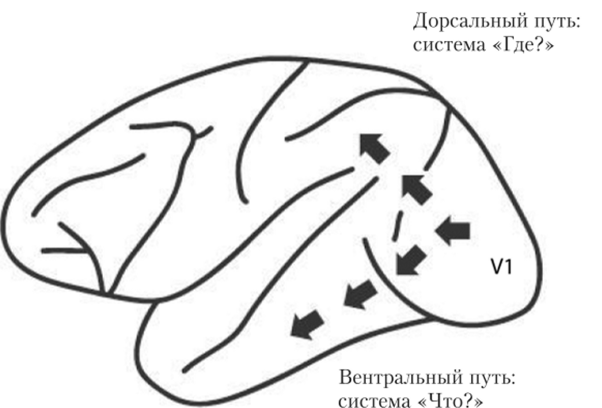
Рис. 8.22. Дорсальный и вентральный потоки зрительных путей.
Открытие Анджерлейдер и Мишкина вызвало большой резонанс в нейрофизиологии сенсорно-перцептивных процессов и повлекло за собой дальнейшие детальные исследования. Дэвид Милнер и Мелвин Гудэйл в 1992 г. подвергли критике теорию Мишкина и Анджерлейдер, и вместо двух зрительных систем «Что?» и «Где?» постулировали наличие двух зрительно-моторных систем «Что?» и «Как?» Вентральный поток «Что?» — это система, которая участвует в распознавании не только цвета, формы, текстуры, но и ориентации, наклона и т. д., тем не менее это пассивная система. А для дорсального потока авторы предлагают изменить название с системы «Где?» на систему «Как?», так как этот поток определяет, КАК субъект осуществляет активное действие (с учетом, разумеется, пространственных характеристик объекта)1. Фактами, подтверждающими такую специализацию высших зрительных путей, являются данные о нарушения в деятельности мозга. Так, Гудэйл и Милнер описали случай Д. Ф., женщины 35 лет с поражением латеральной затылочной и затылочновисочной областей, анатомически соответствующих вентральному потоку. В результате такого поражения пациентка утратила способность соотносить наклон плоского объекта по отношению к по-разному ориентированным отверстиям. Однако, когда ее попросили вставить карту в отверстие (например, как мы вставляем письмо в почтовый ящик) (рис. 8.23), то она справилась с этим заданием при любой ориентации отверстия[4][5][6] (результаты эксперимента с Д. Ф. и контрольным (здоровым) испытуемым показаны на рис. 8.24). Такой результат показывает, что у пациентки нарушена пассивная система восприятия ориентации, но сохранена система активных действий. В пользу существования систем «Что?» и «Как?» свидетельствует много экспериментальных фактов1 2.[7][8][9]
Рис. 8.23. Задача на ориентацию в опытах Гудэйла и Милнера2.
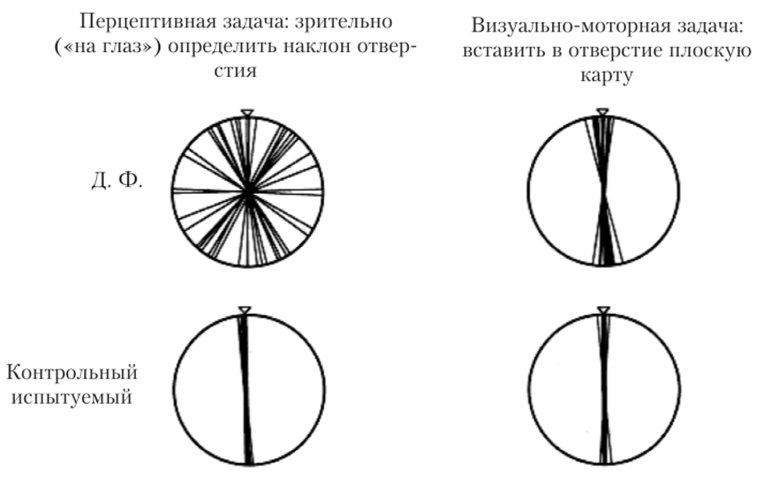
Рис. 8.24. Результат опытов на пассивное восприятие вертикальной ориентации отверстия и активные действия с учетом ориентации отверстия у здоровых испытуемых и пациента с дисфункцией латеральной затылочной и затылочно-височной областей3.
Нейрофизиологические данные о потоках в обработке зрительной информации свидетельствуют в пользу того факта, что распознавание зрительного паттерна происходит одновременно в нескольких направлениях — от выделения в нем простейших элементов (определяющих контуры объекта, а следовательно и его форму) до целостного — структурного — сличения. В пользу последнего свидетельствуют данные о модульной организации коры головного мозга, где модуль — это нейронная структура, специализированная для обработки информации какого-то конкретного типа.
Нейрофизиологические данные показывают, что в коре располагаются относительно независимые структуры, ответственные за восприятие конкретной зрительной информации — не просто параметры стимуляции, а целостные образы (лица, места и т. д.). Например, в исследовании Эдмунда Роллса были измерены ответы нейронов нижневисочной доли коры макаки при предъявлении животному изображения лиц и других типов изображений (полос, абстрактных кривых и т. д.). Ролле и его коллеги в многочисленных экспериментах обнаружили высокую избирательность нейронов именно к изображению лиц1. В этой же области были обнаружены нейроны, чувствительные исключительно к изображению головы. У людей высокой избирательностью к изображениям лиц обладают нейроны веретенообразной извилины, расположенной на нижней поверхности височной доли головного мозга на границе с затылочной долей (рис. 8.25). Обнаружившие такую чувствительность Нэнси Кэнвишер и ее коллеги назвали эту область веретенообразной лицевой зоной (англ. FFA от Fusifomi Face Area)[10][11].
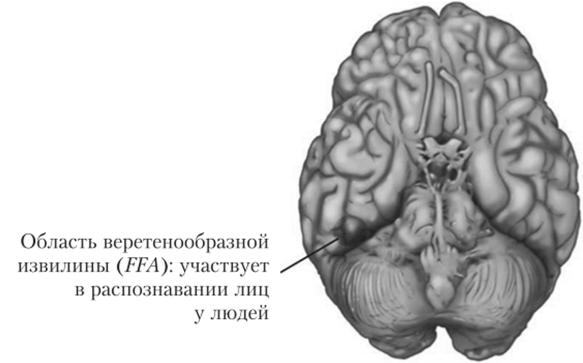
Рис. 8.25. Активность веретенообразной извилины, расположенной на нижней поверхности височной доли головного мозга на границе с затылочной долей, при восприятии лиц.
В одном из исследований испытуемым показывали ряд картинок: на одних были изображены лица, на других — дома. Лица отличались чертами, а дома — расположением окон, дверей и прочих деталей. Испытуемым давали возможность рассмотреть изображения, регистрируя тем временем активность их мозга с помощью фМРТ. Веретенообразная извилина активизировалась, только когда испытуемый рассматривал лица, и оставалась «равнодушной» к домам. Дополнительно в ходе эксперимента ученые обнаружили, что головной мозг распознает лицо в целом, а не отдельные его черты[12][13]. В дальнейшем исследователи обнаружили участки мозга, избирательно чувствительные к изображению человеческого тела или его частей, а также к изображениям местности. В частности, парагиппокампальная область активизируется при восприятии различных сцен, также была получена активность экстрастриарной области при восприятии тела и его частей (рис. 8.26). Такие физиологические данные свидетельствуют, что хотя бы на высших уровнях деятельности зрительной системы имеет место целостное — структурное — распознавание.

Рис. 8.26. Активность парагиппокампальной (РРА) (а) и экстрастриарной (ЕВА) (б) областей при рассматривании разного рода стимулов:
нижняя строка образов для каждой области — набор объектов, не вызывающих активности зоны2
- [1] Mishkin М., Ungerleider L. G. Two Cortical Visual Systems // Ingle D.J., Goodale M. A.(Eds.) Analysis of Visual Behavior. Cambridge: The MIT Press, 1982. P. 549—586.
- [2] Цит. по: Mishkin М., Ungerleider L. G., Macko К. Object vision and spatial vision: Twocortical pathways // TINS. 1983. Vol. 6. P. 414—417.
- [3] Cm.: Distler C., Boussaoud D., Desimone R., Ungerleider L. G. Cortical connections of inferiortemporal area TEO in macaque monkeys //J. Comp. Neurol. 1993. Vol. 334. P. 125—150, а также: Pessoa L., Ungerleider L. G. Top-down mechanisms for working memory and attentional 3
- [4] 3 processes // Gazzaniga М. S. (Ed.) The New Cognitive Neurosciences, MIT Press, 2004. P. 919—930.
- [5] Goodale M. A, Milner A. D. Separate visual pathways for perception and action // TrendsNcurosci. 1992. Vol. 15 (1). P. 20−25.
- [6] Goodale M. A., Milner D. et al. A neurological dissociation between perceiving objects andgrasping them //Nature. 1991. Vol. 349. P. 154—155.
- [7] Например, см.: Milner D., Dyde R. Two illusions of perceived orientation: one fools all of thepeople some of the time; the other fools all of the people all of the time // Exp Brain Res. 2002.Vol. 144 (4). P. 518−527.
- [8] Цит. по: Фрит К. Мозг и душа. Как нервная деятельность формирует наш внутренниймир. М.: Corpus, Астрсль, 2012.
- [9] Goodale М. А., Humphrey G. К. The objects of action and perception // Cognition. 1998.Vol. 67(1−2). P. 181−207.
- [10] См.: Rolls Е. Т. Neurons in the cortex of the temporal lobe and in the amygdala of themonkey with responses selective for faces // Human Neurobiol. 1984. Vol. 3. P. 209—222, а также: Rolls E. T. Neurophysiological mechanisms underlying face processing within and beyond thetemporal cortical visual areas // Phil. Trans: Biol. Sci. 1992. Vol. 335 (1273). P. 11—21.
- [11] KanwisherN. etal. The Fusiform face area: a module in human extrastriate cortex specializedfor face perception //The Journal of Neuroscience. 1997. 17 (11): 4302—4311.
- [12] Yovel G. r Kanwisher N. Face perception: domain specific, not process specific // Neuron.2004. Vol. 44 (5). P. 889−898.
- [13] Kanwisher N. The ventral visual object pathway in humans: evidence from fMRI // Chalupa L.yWernerJ. (Eds.) The Visual Neurosciences. Cambridge, MA: MIT Press, 2003. P. 1179—1189.