Постлактационная инволюция молочной железы

Кроме того, из-за уменьшения потребления молока ребенком в альвеолах долек накапливается FIL (см. подпараграф 4.2.2), который будет тормозить образование молока. Происходит это в первую очередь, по-видимому, в альвеолах долей, из которых ребенок менее эффективно выводит молоко. Например, молочный ход в соске, через который происходит выход молока из доли, имеет наименьший диаметр и… Читать ещё >
Постлактационная инволюция молочной железы (реферат, курсовая, диплом, контрольная)
Всемирная организация здравоохранения рекомендует кормящим грудью женщинам вводить с 5—6 месяцев прикорм, при общей продолжительности грудного вскармливания 1,5—2 года. Объем прикорма увеличивается со временем так, что потребность в молоке для ребенка уменьшается. В связи с этим, как показывают многочисленные обследования, количество секретируемого молочными железами молока также снижается. На рис. 4.14, а в качестве примера представлен график ежесуточной продукции молока и объема молочных желез женщин, которые начали прикорм детей с шести месяцев после родов (Kent et al., 1999). В течении первых шести месяцев после родов продуктивность практически не изменялась, а затем происходило уменьшение количества секретируемого молока и объема молочных желез. Снижение продуктивности молочной железы, вероятно, происходило по нескольким причинам. Общей причиной снижения продуктивности для всех долей молочной железы являлось, по-видимому, сокращение количества кормлении и времени кормлении, в результате чего уменьшался выход из аденогипофиза гормона пролактина — главного гормона у женщин, необходимого для воспроизводства молока (см. подпункт 3.2.4.3.4).
Кроме того, из-за уменьшения потребления молока ребенком в альвеолах долек накапливается FIL (см. подпараграф 4.2.2), который будет тормозить образование молока. Происходит это в первую очередь, по-видимому, в альвеолах долей, из которых ребенок менее эффективно выводит молоко. Например, молочный ход в соске, через который происходит выход молока из доли, имеет наименьший диаметр и, соответственно, высокое сопротивление движению молока (см. пункт 2.1.3.2). Поэтому ребенок будет в первую очередь опорожнять доли, чьи молочные ходы в соске обладают большим диаметром. Альвеолы долей, из которых не выводится молоко, будут переполняться молоком, в результате чего в них начнется процесс постлактационной инволюции. Термин «инволюция» взят из латинского языка для обозначения обратного развития молочной железы (involutio — свертывание). К сожалению, процессы постлактационной инволюции молочной железы женщины на клеточном и молекулярном уровнях остаются не изученными. Наиболее исследованными к настоящему времени среди различных млекопитающих являются клеточные и молекулярные механизмы постлактационной инволюции у молочных желез мышей, благодаря использованию методов генной инженерии (Watson, 2006а, b; Watson, Kreuzaler, 2011). У мышей в соске каждой молочной железы, а их у нее 10, имеется один проток, который неоднократно делится до тончайших веточек, образуя альвеолы. Множественные молочные железы у мышей функционируют независимо друг от друга. У женщин каждая молочная доля имеет свой проток, который открывается на кончике соска. Количество долей в одной молочной железе женщин варьирует от 5 до 12 (King, Love, 2006) (см. подпараграф 2.1.3), и они также функционируют независимо друг от друга. При изучении постлактационной инволюции у молочных желез лактирующих мышей используют блокирование выхода молока из железы с помощью заклеивания выхода протока соска какой-либо молочной железы (Li et al., 1997). В результате из нее молоко детенышами не выводится, в то время как из других желез детеныши могут беспрепятственно извлекать молоко. Данная ситуация в какой-то мере напоминает описанное выше торможение выведения молока из доли молочной железы женщины с тонким протоком в соске. Здесь надо отметить, что мышь кормит детенышей довольно часто и средний интервал между кормлениями составляет 35 ± 2 мин. (Толкунов, Марков, 2005). Увеличение интервала между кормлениями вызывает переполнение альвеол молочных желез. Полагают, что стазис протоков и переполнение молоком альвеол и отходящих от них протоков являются первичным сигналом для начала инволюции (Li et al., 1997), во время которой происходит апоптоз секреторной ткани. Поскольку данные о постлактационнои инволюции молочной железы женщины на клеточном и молекулярном уровнях практически отсутствуют, на рис. 4.14, б в качестве примера представлена кратко схема инволюции альвеолы молочной железы мыши (Watson, 2006b). В процессе инволюции молочных желез мышей выделяют две фазы (см. рис. 4.14, б). В первые 6—12 ч начальной фазы в результате блокировки выхода молока во внутриальвеолярном объеме увеличивается концентрация специфических внеклеточных лигандов и цитокинов, таких как трансформирующий фактор роста (33 (TGF-(33), фактор некроза опухолей a (TNFa), лейкемия-ингибирующий фактор (LIF). Эти агенты индуцируют процесс апоптоза через активацию рецепторов клеточной гибели (рецепторов смерти) (первый путь) и активацию проапоптозного транскрипционного фактора STAT3 (второй путь). Умершие клетки заталкиваются в полость альвеолы. При этом нормально функционирующие соседние клетки двигаются друг к другу и смыкаются латеральными поверхностями, сохраняя тем самым в течение некоторого времени целостность альвеолы. Первая фаза инволюции является обратимой. В том случае, если через 48 ч сосок расклеивается и детеныш начинает выводить молоко, функция данной молочной железы возобновляется (Watson, 2006b). Вслед за обратимой первой фазой инволюции после 48 ч отсутствия выведения молока начинается вторая фаза инволюции (см. рис. 4.14, б).


а — ежесуточный объем молока и объем молочных желез (по Kent et al., 1999). На графиках под каждой точкой в скобках отмечено количество наблюдений; б — схематическое изображение этапов инволюции молочной альвеолы мыши (по Watson, 2006а): 1 — нормально функционирующая секреторная клетка,.
2 — мертвая секреторная клетка, 3 —клетка, измененная в результате апоптоза, 4 — рецептор клеточной гибели, 5 — лиганды рецепторов клеточной гибели, 6 — адипоцит, 7 — молочные глобулы, 8,9 — лейкемия-ингибирующий фактор (LIF). Цифрами обозначено время инволюции в часах В результате увеличения в экстраклеточной среде концентрации и активности таких ферментов, как матричные металлопротеиназы (ММР), происходит разрушение экстраклеточного матрикса вокруг альвеол, что усиливает апоптоз и в финале происходит коллапс альвеол. Одновременно осуществляется дифференциация окружающих альвеолу адипоцитов так, что они начинают вновь заполняться жиром. В этой фазе наблюдаются процессы аутофагии и гетерофагии, т. е. самопереваривания тканей молочной железы. Растет количество ферментов лизосом — активных органелл, переваривающих клеточные останки. Вторая фаза инволюции является необратимой, поэтому альвеолы, в которых она началась, уже не могут вновь лактировать. Нужно чтобы снова начались процессы дифференцировки и пролиферации железистой ткани, как это происходит во время беременности. Следует отметить, что степень различия или сходства в молекулярных и клеточных механизмах между инволюцией молочной железы мыши и женщины неизвестна. Тем не менее в некоторых научно-популярных обзорах инволюция молочных желез мышей и женщин представляется идентичной. Вместе с тем имеющиеся экспериментальные данные указывают, что, например, временные параметры инволюции молочных желез мышей и женщин существенно различаются. Так, при резкой остановке кормлений ребенка процесс инволюции у женщины занимает до 42 суток (Hartmann, Kulski, 1978), в то время как у мышей 6—7 суток (Watson, 2006а), т. е. в 6—7 раз дольше, чем у мышей. Возможно, одной из причин является более медленная скорость инволюционных процессов, которая обусловлена особенностями лактации женщины. В частности, промежуток между кормлениями во время установившей лактации женщины составляет 2,5—3 ч, а у мышей 0,5 ч. Таким образом, время накопления молока для очередного кормления ребенка в 5—6 раз больше, чем у мыши для кормления детенышей. Соответственно, по-видимому, при непредвиденных обстоятельствах при длительной задержке кормления ребенка, чтобы не начиналась инволюция железистой ткани, время начала инволюции у женщины сдвинуто на более поздний срок. Процесс постлактационной инволюции молочной железы у женщины в норме, т. е. при введении прикорма ребенку и одновременном сниженном потреблении ребенком молока развивается плавно и занимает сравнительно большой промежуток времени (см. рис. 4.14, а). Причем отдельные доли молочных желез подвергаются инволюции асинхронно. Это легко определяется по количеству струек молока при сжатии ареолы пальцами. Так, в начале инволюции сжатие ареолы молочной железы женщины сопровождается появлением в среднем 5—10 струек молока. В дальнейшем количество струек начинает уменьшаться, что свидетельствует о прекращении образования молока в некоторых долях.
Вместе с тем, как уже упоминалось выше, при особых условиях (болезнь ребенка или матери) женщина вынуждена резко прекратить кормление. В этом случае процесс инволюции развивается значительно быстрее и может закончится в течение 1—1,5 месяца.
Помимо уменьшения общего количества секретируемого молока при «медленной» и «быстрой» инволюции молочной железы происходит изменение концентрации различных компонентов молока.
Введение
прикорма и начало инволюции дополнительно вызывает ускорение изменений в составе молока. К сожалению, о влиянии инволюции молочной железы на содержание различных компонентов молока в литературе имеются единичные работы, касающиеся динамики концентраций лишь некоторых компонентов молока. Так, при изучении состава молока у женщин при введении прикорма детям возраста 5—7 месяцев (Garza et al., 1983) было обнаружено, что происходит в течение последующих трех месяцев постепенное изменение концентраций некоторых компонентов: увеличение концентраций белка от 135 ± 6 до 193 ±21 MrN/ЮО мл; концентрация жира повышалась от 3,6 ± 0,8 до 7,5 г/100 мл в течение первых 10 недель, но затем уменьшалось до 4,07 ± 0,7; железа — от 29 ± 6 до 50 ± 5 мкг/1000 мл; натрия — от 136 ± 16 до 297 ± 57 мг/1000 мл; концентрация кальция не изменялась; цинка уменьшалась — от 1,2 ± 0,2 до 0,7 ± 0,1 мг/1000 мл. В последующих работах (Perrin et al., 2016) были проведены исследования динамики концентраций ряда компонентов молока женщин, кормивших детей в течении 11—17 месяцев лактации.
Дети наряду с молоком получали прикорм, т. е. молочные железы этих женщин подвергались медленной инволюции. Оказалось, что в этот период происходило увеличение кроме концентрации общего белка от.
- 1.6 ± 0,2 до 1,8 ± 0,3 г/100 мл; белков, которые обладают бактерицидными свойствами, и в частности лактоферрина, — от 180 ± 66 до 280 ± 180 мг/100 мл; лизоцима — от 58 000 ± 31 000 ед/мл; IgA — от 21 ±
- 7.7 до 29 ± 10 мг/100 мл. Происходило также повышение общего содержания натрия от 70 ± 19 до 86 ± 35 мкг/мл и одновременно снижение общей концентрации цинка от 560 ± 339 до 420 ± 310 нг/мл и кальция от 200 ± 29 до 180 ± 30 мкг/мл. Не было обнаружено изменений в этот период концентрации лактозы, жира, железа и калия.
В литературе имеется одна работа (Hartmann, Kulski, 1978), в которой была исследована динамика ряда минеральных и органических веществ в молоке при «быстрой» инволюции молочных желез женщин. На рис. 4.15 представлены графики изменения концентрации различных компонентов молока у женщин 251—443 дня лактации.
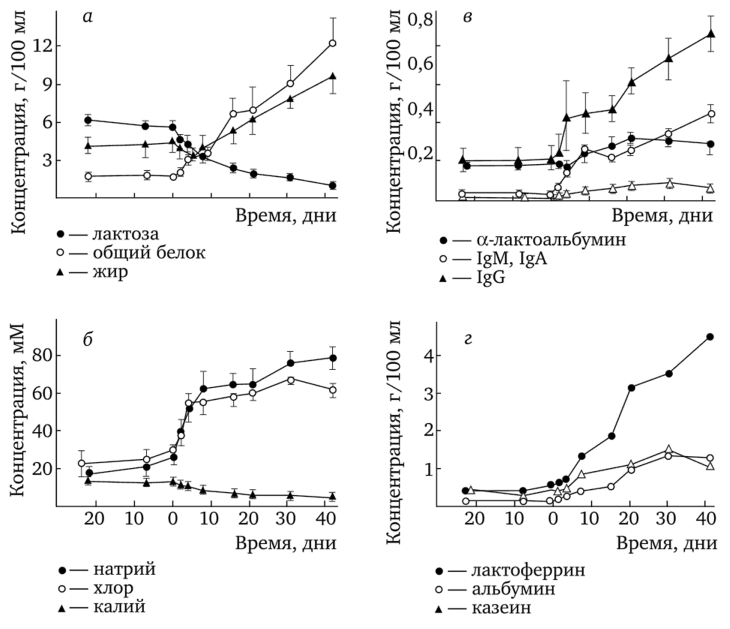
Рис. 4.15. Изменение концентрации некоторых компонентов молока при резкой остановке кормлений ребенка (по Hartmann, Kulski, 1978):
У женщин в определенный момент останавливали грудное вскармливание так, что молоко из желез не выводилось за исключением небольших проб (0,5—5,0 мл), необходимых для анализа В течение 45 дней количество молока быстро снижалась. Сравнение данных изменений концентрации компонентов молока при «медленной» и «быстрой» инволюции показывает в большинстве случаев однонаправленность этих изменений. Здесь надо отметить, что ряд компонентов молока, которые анализировались при «быстрой» инволюции, не определялись в молоке при «медленной» инволюции и наоборот. При «быстрой» инволюции концентрации общего белка, жира, натрия и хлора увеличивались, концентрации калия и лактозы снижались (см. рис. 4.15, а, б). Представленные данные позволяют считать, что во время инволюции происходит увеличения проницаемости плотных межклеточных контактов к натрию, хлору, калию, лактозе. В результате начинает происходить выравнивание концентраций данных компонентов молока между экстраклеточной средой и внутриальвеолярным объемом. То есть ситуация, обратная переходу молочных желез от беременности к лактации, при которой, как полагают, происходит блокирование проницаемости плотных контактов на вторые-третьи сутки после родов (см. параграф 3.1). Характерным для состава молока при «медленной» и «быстрой» инволюции является заметное повышение белков, обладающих антимикробными свойствами, таких как а-лактальбумин, лизоцим, лактоферрин, иммуноглобулины А, М, G (Hartmann, Kulski, 1978; Perrin et al., 2016), (см. рис. 4.15, в, г). Эти белки важны в первую очередь для женщины, поскольку стазис молока и переполнение альвеолярно-протоковой системы молоком при инволюции способствуют росту концентрации бактерий в молоке и развитию воспалительных процессов. Как показывает клиническая практика, в настоящее время для устранения возможных болезненных нагрубаний некоторых долей молочных желез, когда женщина после сравнительно большого срока кормления ребенка с прикормом (12—14 месяцев) резко прекращает кормить ребенка, дополнительно используется агонист дофамина — каберголин (достинекс). После сцеживания нагрубшей доли на фоне действия достинекса секреция молока в этой доле в дальнейшем блокируется. Кроме того, в недавних исследованиях, правда, на молочных железах коров (Boutinaud et al, 2017), было обнаружено, что в период инволюции молочных желез каберголин наряду с блокированием выхода пролактина усиливает отслаивание эпителиальных клеток альвеол и активирует матричные металлопротеиназы ММР2 и ММР9, т. е. ускоряет вторую фазу инволюции (см. рис. 4.14, б). Не исключена возможность, что каберголин ускоряет апоптоз и в альвеолах молочных желез женщин.
Таким образом, в результате постлактационной инволюции структура и функция молочных желез возвращаются к состоянию, которое характерно для женщин с нормальным менструальным циклом. При этом молочные железы готовы снова начать лактировать в случае наступления очередной беременности и родов.